have been retained. The official nomenclature for structures of the kidney has been established by the Renal Commission of the International Union of Physiological Sciences (12). This terminology is used in this chapter (Table 1.1).
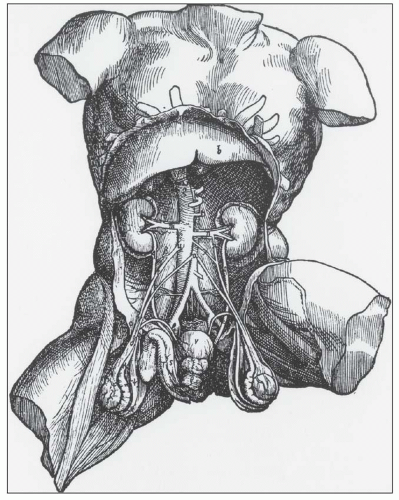 FIGURE 1.1 Andreas Versalius’ 1543 illustration of the male genitourinary tract. Although most anatomic details are correct, the left kidney is incorrectly placed lower than the right kidney. (From Book 5 De Humani Corpus Fabrica.) |
is a normal anatomic variant without functional consequences (22,23,24). In some of the kidneys, three subtle zones are created by two shallow superficial grooves that radiate from the hilum to the lateral border. The three regions correspond roughly to the upper pole, middle zone, and lower pole and usually reflect regions drained by the segmental veins.
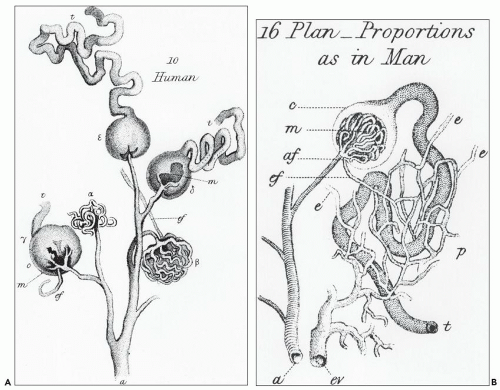 FIGURE 1.2 William Bowman’s 1842 dye study. A: Illustration of the glomerular vascular supply. B: Illustration of the relationship between the efferent arteriole and peritubular capillary plexus and the proximal tubule. (From Bowman W. Philos Trans R Soc Lond Biol 1842;132:57.) |
ducts of Bellini, creating the potential for intrarenal reflux during urinary tract obstruction and infection (30,31).
TABLE 1.1 Nomenclature of the kidney | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
collecting ducts (CDs). The papilla protrudes into a minor calyx. Its tip, the area cribrosa (with cribriform appearance grossly), has from 20 to 70 openings of the papillary CDs (Bellini ducts); the large variation reflects simple versus compound pyramid arrangement (24,26,28).
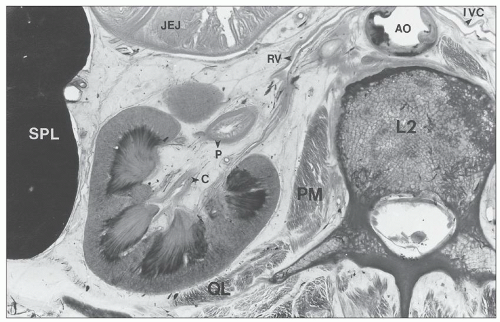 FIGURE 1.3 Cross section of a human cadaver at the level of the second lumbar vertebra viewed from above, demonstrating the relationships between the left kidney and adjacent organs. L2, second lumbar vertebra; AO, aorta; PM, psoas muscle; QL, quadratus lumborum muscle; SPL, spleen; JEJ, jejunum; C, calyx; P, pelvis; RV, renal vein; IVC, inferior vena cava. |
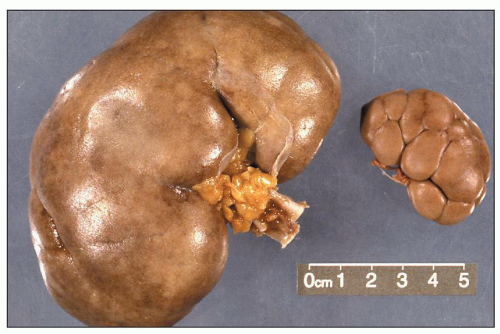 FIGURE 1.4 Composite adult and newborn human kidneys. Notice the smooth subcapsular surface of the adult kidney and the prominent lobations of the newborn kidney. |
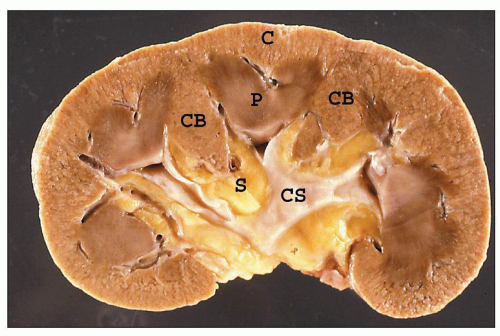 FIGURE 1.5 A hemisected human kidney shows the cortex (C) with columns of Bertin (CB) and renal pyramids (P). The renal papillae protrude into the minor calyces, which unite to form the major calyces. The collecting system (CS) is nestled within the fatty tissue of the renal sinus (S). |
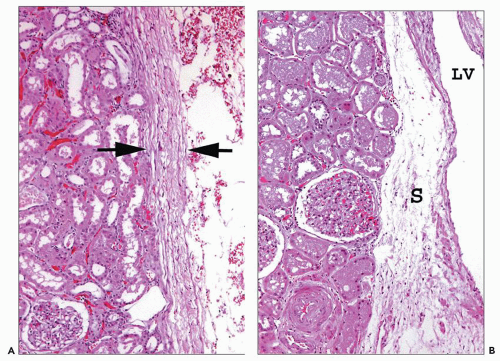 FIGURE 1.6 Human kidney. A: A dense connective tissue capsule (between arrows) separates the peripheral cortex from the perinephric fat (×100). B: The renal tubules and interstitium of the columns of Bertin have no connective tissue separation from the sinus (S) and its abundant lymphatics and vessels. (×120.) LV, lobar vein. |
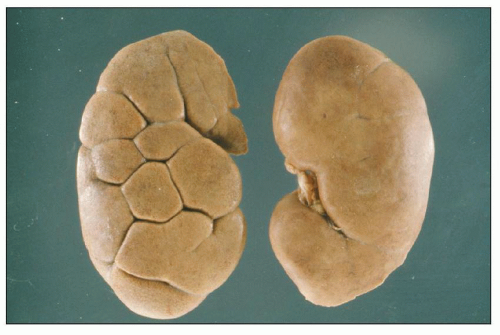 FIGURE 1.7 Two normal adult human kidneys, one with retained lobations. |
either anterior or posterior branches or arising separately from the aorta as an aberrant, accessory, or polar artery (Figs. 1.13 and 1.14), which occurs in 25% of the kidneys (32). Another variation is supply of the upper or lower pole by the posterior segmental artery or a branch from the posterior segmental artery.
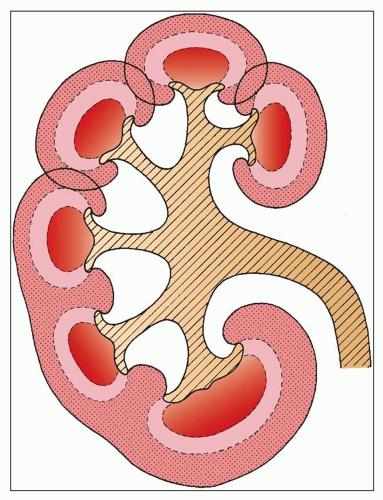 FIGURE 1.8 Diagram of the arrangement of the cortex and medulla in the human kidney and of its possible development. Unipapillary kidney anlagen fuse to form a multipapillary kidney (upper part of the drawing). The interpyramidal cortical intrusions (septa of Bertin) reach down to the renal sinus. In addition to the fusion of the cortical tissue, pyramidal fusion occurs and is generally found in the polar regions (compound papilla, shown in the lower part of the drawing). |
afferent arteriole (Figs. 1.15 and 1.16). The efferent arterioles, on exiting the glomeruli, form a portal system of capillaries that supplies the adjacent cortical tubules, or provides the main arteriolar flow to the renal medulla in the case of juxtamedullary glomeruli (Figs. 1.2B and 1.17; see Cortical Microvascularization section below). A few interlobular arteries reach the renal capsule and anastomose with branches of the suprarenal and gonadal arteries.
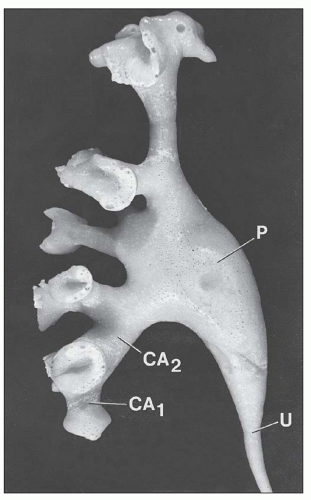 FIGURE 1.9 Minor calyces (CA1) fuse to form major calyces (CA2) that finally become the renal pelvis (P), which tapers to continue on as the ureter (U). Most calyces are angled toward the renal pelvis. Corrosion cast of a human renal pelvis. (From Sampaio FJB, Mandarim-De-Lacerda CA. 3-Dimensional and radiological pelvicaliceal anatomy for endourology. J Urol 1988;140:1352.) |
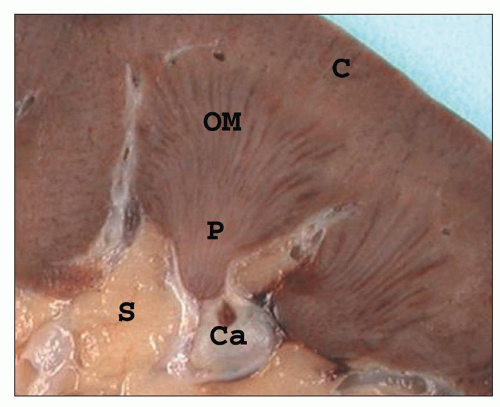 FIGURE 1.10 A simple human renal pyramid with a convex papilla (P) nestled within a minor calyx. S, sinus; OM, outer medulla; C, cortex; Ca, calyx. |
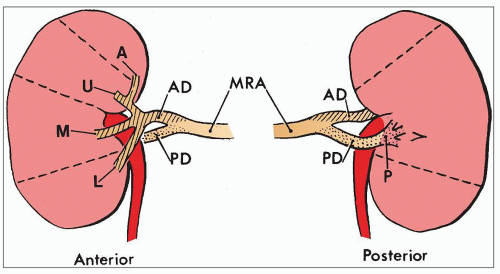 FIGURE 1.11 Diagram of the most common pattern of arterial supply to the kidneys demonstrating the main renal artery, anterior and posterior branches, and five segmental arteries. MRA, main renal artery; PD, posterior division; AD, anterior division. Segmental arteries are indicated by A (apical), U (upper), M (middle), L (lower), P (posterior). (Modified from Graves FT. The anatomy of the intrarenal arteries and its application to segmental resection of the kidney. Br J Surg 1954;42:132.) |
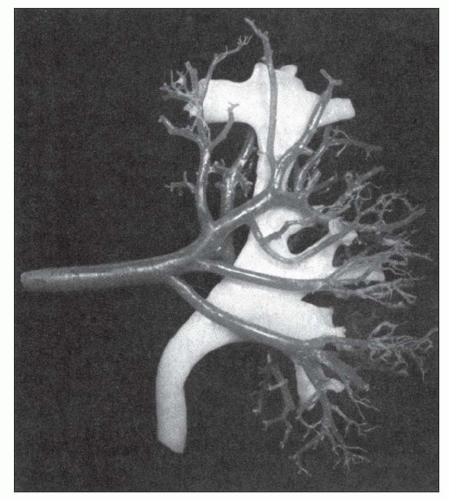 FIGURE 1.12 Cast of the arterial supply and collecting system of a human kidney. Notice the four anterior segmental arteries. (From Sampaio FJB, Aragao AHM. Anatomic relationship between the intrarenal arteries and the kidney collecting system. J Urol 1990;143:679.) |
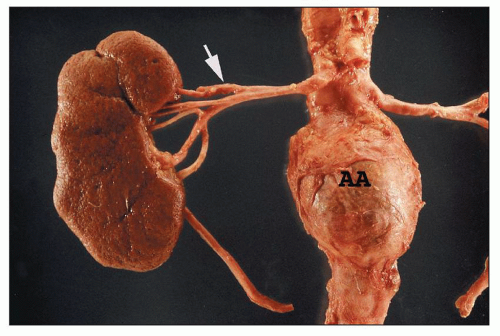 FIGURE 1.13 Human right kidney and aorta. The aorta has an aneurysm (AA). The main renal artery and its five segmental arteries are visible although not in the most common arrangement. The upper segmental artery branches first (arrow), and the posterior segmental artery appears to arise from the anterior group. |
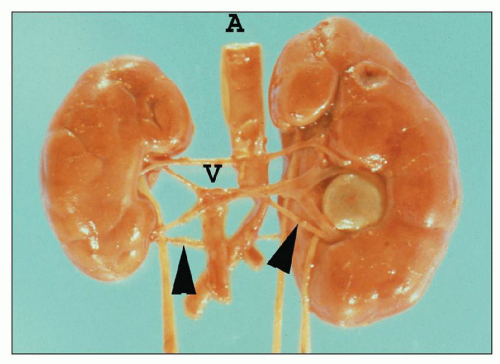 FIGURE 1.14 Newborn human kidneys. There are bilateral lower pole accessory (segmental) arteries (arrowheads). The crossing polar artery and segmental vein of the nonrotated duplex (two ureters) left kidney produced ureteropelvic junction obstruction. A, aorta; V, vena cava. |
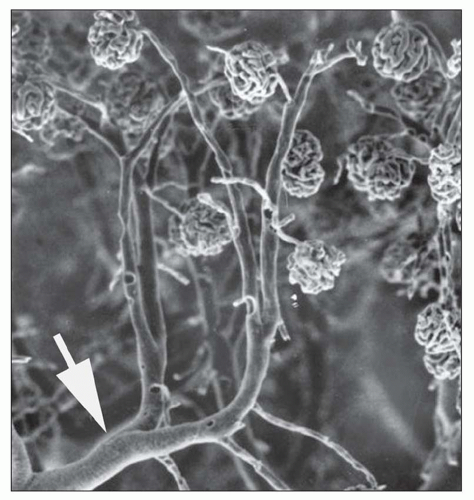 FIGURE 1.15 Arterial cast of the rat kidney. Two interlobular arteries give rise to arterioles that supply a single glomerulus. Arrow, arcuate artery. (From Gattone II VH, Evan AP, Willis LR, et al. Renal afferent arteriole in the spontaneously hypertensive rat. Hypertension 1983;5:8.) |
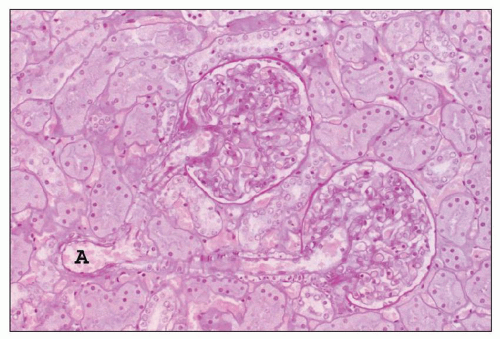 FIGURE 1.16 Normal human kidney. The interlobular artery (A) gives rise to two arterioles; each supplies one glomerulus. (PAS, ×200.) |
from juxtamedullary glomeruli. Their efferent arterioles course directly into the medulla, forming the descending vasa recta (Fig. 1.17). The second blood supply originates from an interlobar artery as it courses along a minor calyx. Several spiral artery branches enter the papilla at the calyceal fornices, sending arterioles to the papillary tip. These arterioles anastomose freely with arterioles from the opposite side, forming a plexus around the distal ducts of Bellini.
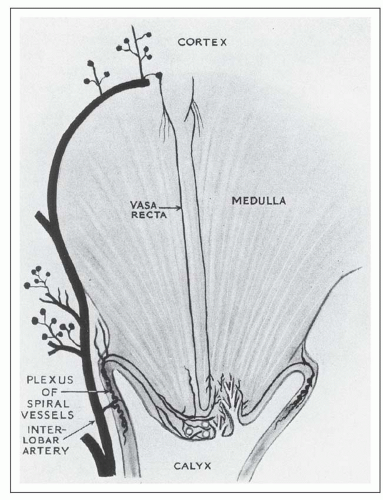 FIGURE 1.17 Diagram of the renal medulla showing its dual blood supply. (From Baker SB. The blood supply of the renal papilla. Br J Urol 1959;31:57. Reprinted from Wiley-Liss, Inc., Wiley Publishing Inc., a subsidiary of John Wiley & Sons, Inc., with permission.) |
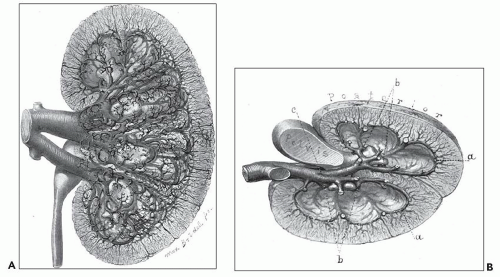 FIGURE 1.18 Brödel’s 1903 artist’s rendition of the human kidney following celloidin injection with tissue digestion, demonstrating details of the lush venous return. A: Anterior view of the left kidney. For the sake of clearness, the small veins of the cortex of the anterior portion have been omitted. B: The transverse section viewed from above. There is no collecting vein posterior to the renal pelvis; all of the veins of the posterior region cross over to the anterior portion between the necks of the minor calices (b) to join the veins of the anterior region at a point indicated by c. (Brödel M. The intrinsic blood vessels of the kidney and their significance in nephrotomy.Johns Hopkins Hosp Bull 1901;118:10.) |
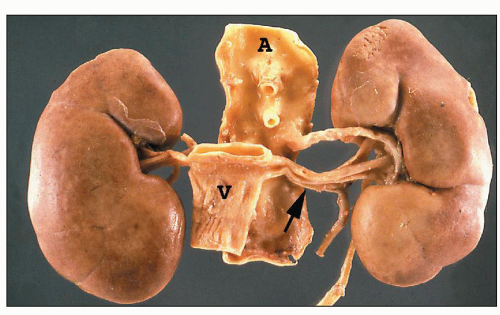 FIGURE 1.19 Human kidneys with aorta (A) and vena cava (V). The left main renal vein (arrow) is much longer than the right main renal vein. |
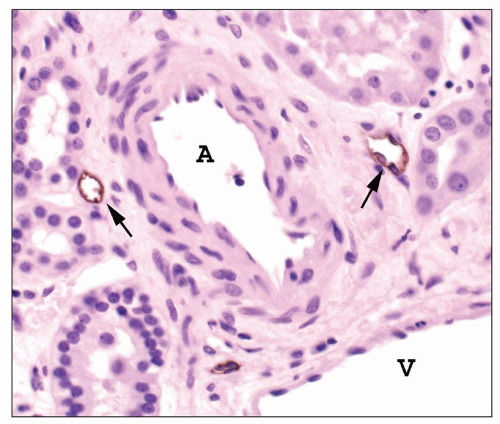 FIGURE 1.20 Human interlobular arteries and lymphatics. Adjacent to an interlobular artery (A) are several small lymphatics (arrows). These will converge and progressively enlarge as they drain toward the medulla. The lymphatic endothelium is stained for podoplanin, a lymphatic endothelial cell marker. V, vein (Immunoperoxidase stain for podoplanin, ×400.) |
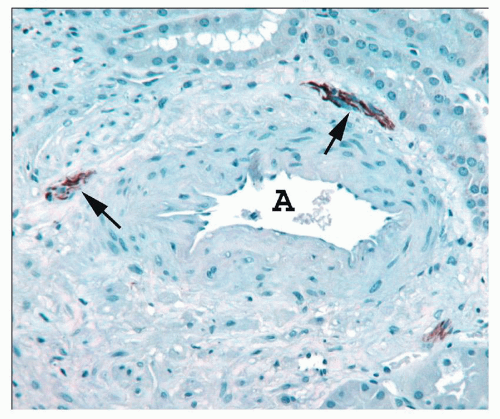 FIGURE 1.21 Human interlobular artery. The adventitia of this artery (A) contains three small nerves stained for neurofilament (arrows). (Immunoperoxidase stain for neurofilament.) |
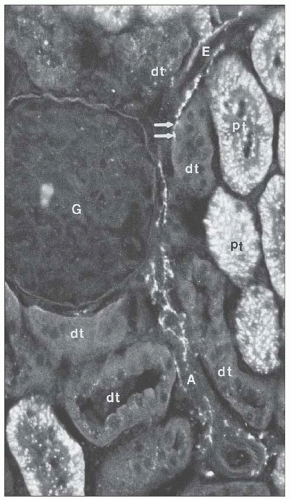 FIGURE 1.22 Nerve fibers viewed by catecholamine fluorescence histochemistry are seen along the afferent (A) and efferent (E) arterioles of the rat kidney. A spot of fluorescence (arrows) appears adjacent to a distal tubule (DT). G, glomerulus; PT, proximal tubule. (×306.) (From Barajas L. Innervation of the renal cortex. Fed Proc 1978;37:1192.) |
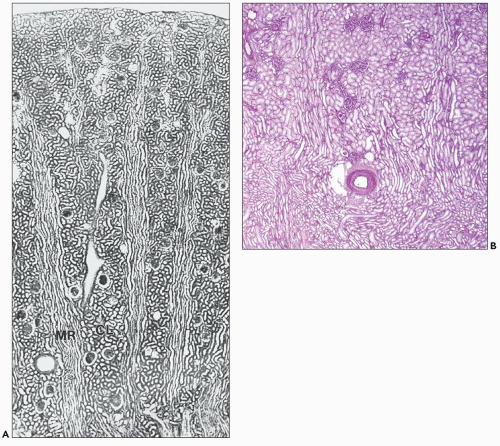 FIGURE 1.23 A: Longitudinal section through the cortex of a human kidney demonstrates the arrangement of the cortical labyrinth (CL) and the medullary rays (MRs). The cortical labyrinth contains the interlobular vessels and the glomeruli together with the convoluted tubules; the medullary rays contain the straight tubular portions and collecting ducts. (Paraffin section, ×75.) B: This section from a human kidney shows two medullary rays with their longitudinally oriented tubules. An artery and glomeruli are centrally located within the cortical labyrinth between the two medullary rays. Notice that the tubules of the outer stripe of the outer medulla located beneath the artery appear similar to those in the medullary rays. (Periodic acid-Schiff, × 20.) |
a loose vascularized connective tissue layer, the lamina propria, with an underlying thin muscularis propria.
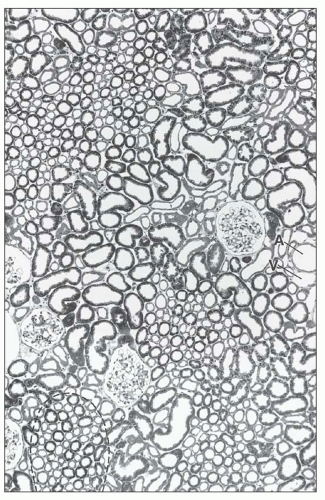 FIGURE 1.24 Cross-section (1-µm section of Epon-embedded tissue) through the cortex of the human kidney. In the cortex, the cortical labyrinth can clearly be delineated from the medullary rays (the cross section of one ray is marked by a dashed line). Within the labyrinth, the interlobular vessels (A, artery; V, vein), the glomeruli, and the convoluted tubular segments are found. The medullary rays contain the straight tubular segments and collecting ducts. (×140.) |
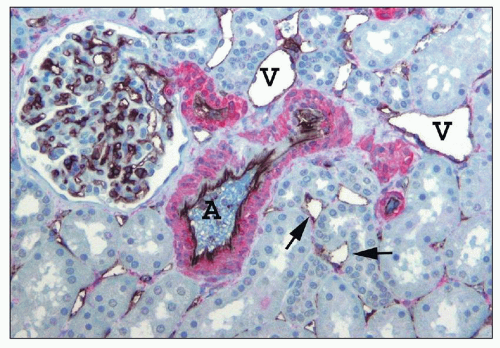 FIGURE 1.25 Human renal cortical labyrinth. The cortical labyrinth contains glomeruli, vessels, and tubules, mostly proximal tubules. The tubules have closely apposed basement membranes with little interstitial space, largely occupied by peritubular capillaries (arrows). The smooth muscle of the interlobular artery (A) and arterioles is stained red for smooth muscle actin. The veins (V) have very thin walls lacking smooth muscle. The endothelial cells of glomeruli, peritubular capillaries (arrows), and larger vessels are stained black for an endothelial marker, CD31. (×150.) |
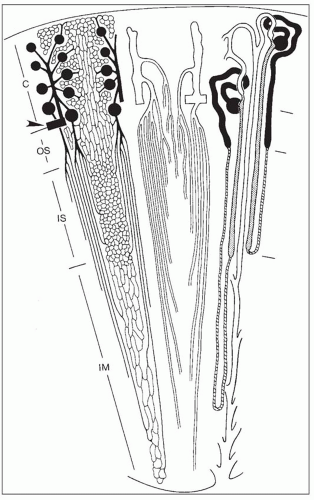 FIGURE 1.26 Schematic diagram of microvasculature and nephrons (not drawn to scale). C, cortex; OS, outer stripe; IS, inner stripe; IM, inner medulla. Left: Arterial vessels and capillaries. An arcuate artery (arrowhead) gives rise to an interlobular artery (cortical radial artery) from which afferent arterioles originate. Efferent arterioles from superficial and midcortical glomeruli split off into the cortical peritubular capillaries. Efferent arterioles of juxtamedullary glomeruli descend into the outer stripe and divide into the descending vasa recta, which supply the different capillary plexuses in the medulla. Middle: Venous vessels. Interlobular veins start in the superficial cortex. Stellate veins (characteristically found in the human kidneys), which begin on the renal surface, are not shown. The deep portions of interlobular veins and arcuate veins accept the ascending vasa recta, which drain the venous blood from the medulla. Ascending vasa recta and descending vasa recta together establish the vascular bundles. Note the dense pattern of ascending recta traversing the outer stripe as wide, tortuous channels. Right: A short- and a long-looped nephron together with a collecting duct. Glomeruli and proximal tubules are drawn black. Thin limbs are hatched; thick limbs are dotted. Distal convoluted tubules, connecting tubules (including an arcade), and collecting ducts are white. This drawing allows for the correlation of the location of tubules and vessels; the left, middle, and right views should be imagined as being superimposed on each other. |
nephrons are short and quickly transition into a peritubular capillary plexus that forms a uniformly distributed anastomosing vascular lattice amid the cortical tubules of the labyrinth. In the medullary rays the capillary plexus assumes a more longitudinal orientation, following the course of the straight tubules.
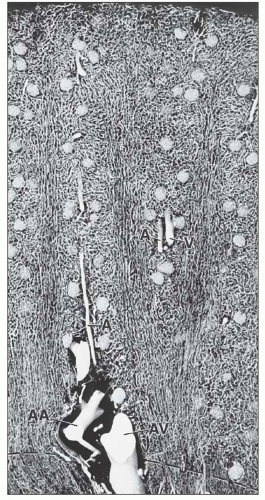 FIGURE 1.27 Longitudinal section through the cortex of a human kidney. The arteries have been injected with silicone rubber (Microfil). The different capillary patterns of the cortical labyrinth and of the medullary rays are visible. Within the cortical labyrinth, the interlobular (corticoradial) arteries (A) and veins (V) as well as the glomeruli are found. Within the medullary rays, the capillary plexus consists of elongated meshes. (×70.) AA, arcuate artery; AV, arcuate vein. |
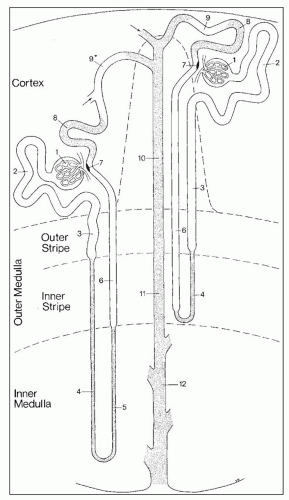 FIGURE 1.28 Schematic drawing of the nephron. This scheme depicts a short-looped and a long-looped nephron together with the collecting system. Not drawn to scale. Within the cortex, a medullary ray is delineated by a dashed line. 1: Renal corpuscle including Bowman capsule and the glomerulus (glomerular tuft). 2: Proximal convoluted tubule. 3: Proximal straight tubule. 4: Descending thin limb. 5: Ascending thin limb. 6: Distal straight tubule (thick ascending limb). 7: Macula densa located within the final portion of the thick ascending limb. 8: Distal convoluted tubule. 9: Connecting tubule. 9*: Connecting tubule of the juxtamedullary nephron that forms an arcade. 10: Cortical collecting duct. 11: Outer medullary collecting duct. 12: Inner medullary collecting duct. (From Kriz W, Bankir L, Bulger RE, et al. A standard nomenclature for structures of the kidney. The Renal Commission of the International Union of Physiological Sciences. Kidney Int 1988;33:1.) |
monkeys, the vascular bundles are of the simple type, and the tubulovascular relationships of the outer stripe are maintained in the inner stripe. The descending thin limbs (DTLs) and ascending thick limbs of the long-looped juxtamedullary nephrons are situated close to the vascular bundle, whereas the DTLs and ascending thick limbs of the short-looped nephron and the CDs are more peripheral (Fig. 1.33). In other species, such as rat, mouse, and desert rodents, the vascular bundles are of the so-called complex type. In this type, DTLs of shortlooped nephrons leave the interbundle region and curve toward the vascular bundles, intermingling with ascending vasa recta. The thick limbs of both short- and long-looped nephrons and the thin limbs of long-looped nephrons remain in the interbundle region with CDs. These histotopographic differences in tubulovascular relationships generally correlate with urinary concentration ability. However, exceptions do occur such as in the hamster with its simple vascular bundles and high concentration capacity.
TABLE 1.2 Tubular segments within each zone of the medulla | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||
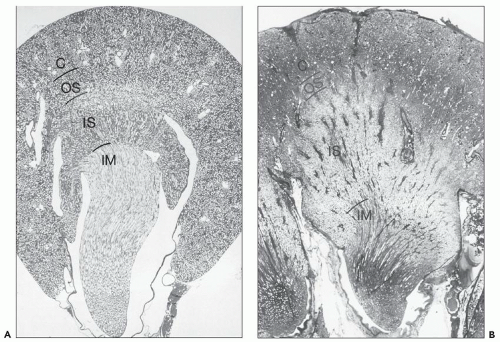 FIGURE 1.29 Longitudinal sections (with respect to the papilla) through a unipapillary kidney (rat) (A) and a single papilla of a multipapillary kidney (human) (B). Note the different proportions of the individual zones. The outer stripe in the human kidney is very narrow, and the inner stripe is thick. (Paraffin sections, A: ×10; B: ×3.) C, cortex; OS, outer stripe; IS, inner stripe; IM, inner medulla. |
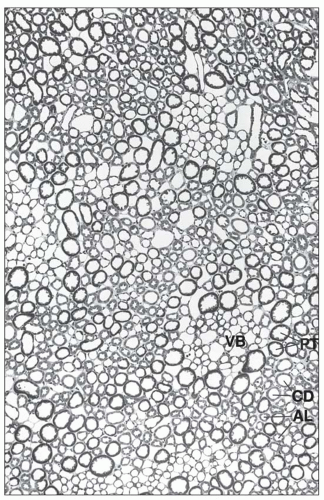 FIGURE 1.30 Cross section (1-µm section of Epon-embedded tissue) through the outer stripe in the outer medulla. The tubules are arranged around the vascular bundles (VB). In the vicinity of the bundles, the nearest segments are the straight proximal and distal tubules of the jux-tamedullary nephrons. More peripherally located to them are the straight proximal and distal tubules of midcortical and superficial nephrons; the collecting ducts are lying distant from the bundles. (×140.) PT, proximal tubule; AL, ascending limb (straight parts of distal tubule); CD, collecting duct. |
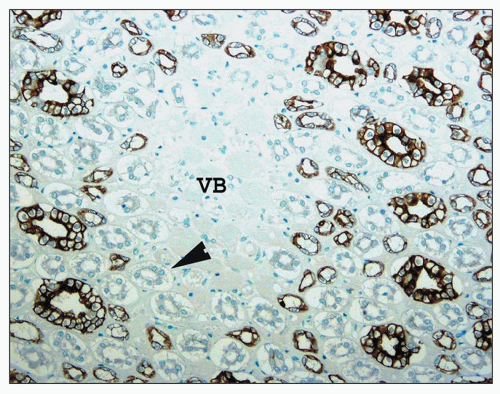 FIGURE 1.31 Human kidney, outer medulla. The central vascular bundles (VB) are surrounded by the descending straight tubules (arrowhead), which are unstained, and the smaller stained ascending thick limb tubules. The largest stained tubules are collecting ducts. (Immunoperoxidase stain for cytokeratin AE 1/3, ×250.) |
which is sparse in true capillaries. They also travel outward from the bundle as they enter the outer stripe. The ascending venules finally empty into interlobular or arcuate veins. The venae rectae and the arteriolae rectae are arranged in close proximity throughout their course as they travel down to the papillary tip and back up. The intermingled venous and arterial limbs compose the countercurrent exchange system that maintains the medullary osmotic gradient.
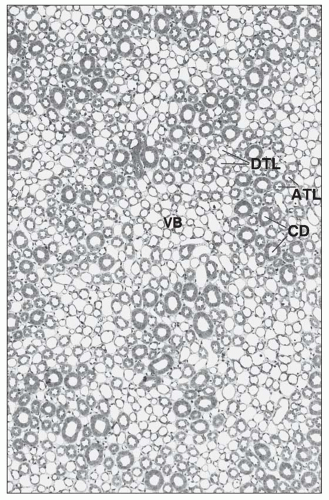 FIGURE 1.32 Cross-section (1-µm section of Epon-embedded tissue) through the inner stripe of the outer medulla. The vascular bundles (VB) are well developed and surrounded by the limbs of short and long loops of Henle in an arrangement similar to that in the outer stripe. (×140.) DTL, descending thin limb; ATL, ascending thick limb; CD, collecting duct. |
The tubular portion of the nephron has complex spatial and topographic relationships with its microvasculature and demonstrates sequential variation in its cellular constitution tightly linked to function.
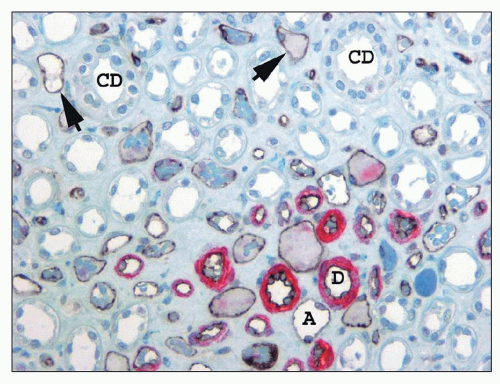 FIGURE 1.33 Human kidney, outer medulla. The descending arteriolae rectae (D) in the vascular bundle have a prominent smooth muscle media (stained red for smooth muscle actin), whereas the thin ascending venae rectae (A) and the interbundle capillary plexus (arrows) have no smooth muscle but can be distinguished from the thin limbs of Henle because their endothelial cell lining is stained black with CD 31. (×350.) CD, collecting duct. |
 FIGURE 1.34 Cross-section (1-µm section of Epon-embedded tissue) through the inner medulla. The vascular bundles (VB) are small and cannot clearly be separated from the surrounding thin limbs (TLs). (×112.) CD, collecting duct. |
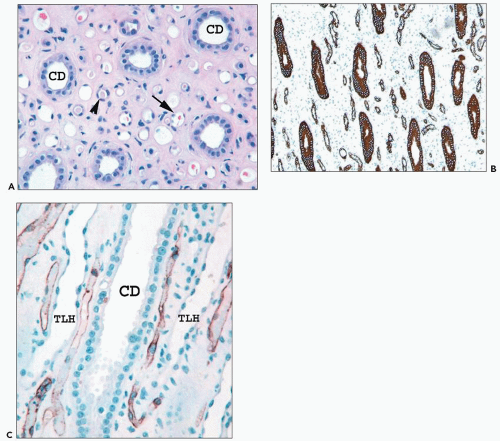 FIGURE 1.35 Human kidney inner medulla. A: The collecting ducts (CDs) are easily identified. However, the thin limbs (arrowhead) and vessels (arrow) appear similar. (×300.) B: The interstitium of the inner medulla is prominent compared with the density of tubules. The thin limbs of Henle and collecting ducts are highlighted by cytokeratin 7 stain. (×300.) C: CD31 labels vascular endothelium of the vasa recta, which are positioned between the thin limb (TLH) and collecting ducts (CDs). The descending vasa recta at the lower right are shown looping back as ascending vasa recta. (×350.) |
the other two, but there is no distinct boundary between these three regions.
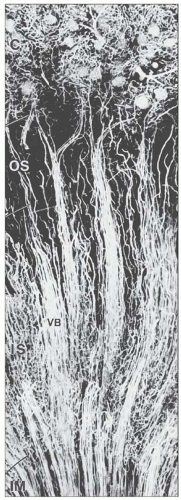 FIGURE 1.36 Arterial vessels of the medulla of a mouse injected with silicone rubber (Microfil). The juxtamedullary glomeruli give rise to efferent arterioles (arrows), which split into the descending vasa recta, establishing the arterial part of the vascular bundle (VB). The bundles are best developed in the inner stripe (IS) and decrease after transition into the inner medulla (IM). C, cortex; OS, outer strip. (×85.) |
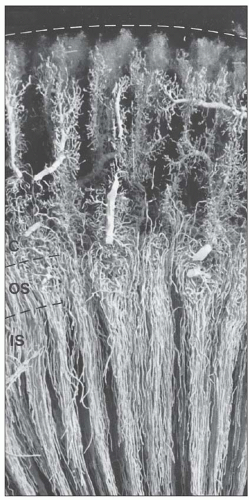 FIGURE 1.37 Venous vessels of rabbit kidney injected with silicone rubber (Microfil). The interlobular veins of the cortex accept the blood from the cortical plexuses and descend to the corticomedullary border. The venous vessels of the medulla are the ascending vasa recta, which ascend within and between the vascular bundles toward the corticomedullary border, where they empty into arcuate veins or the basal portions of the interlobular veins. C, cortex; OS, outer stripe; IS, inner stripe. (×20.) |
this is related to the needs for urinary concentration and water conservation. However, urinary concentration correlates more with the complexity of medullary development rather than simply the number of long-looped nephrons (62,63,64,65,66). In the human kidney, approximately 15% of nephrons are long looped. In the rodent kidney, this is almost doubled to 28%, whereas in cats and dogs, all nephrons are long looped (72).
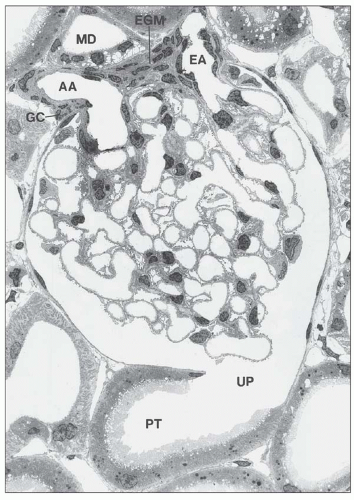 FIGURE 1.38 Glomerulus with juxtaglomerular apparatus of the rat kidney. The afferent arteriole (AA), the efferent (EA), and the extraglomerular mesangium (EGM) are in close contact with the macula densa (MD) of the thick ascending limb. UP, urinary pole; PT, proximal tubule; GC, granular cell. (Electron micrograph, ×900.) |
There are afferent, more peripherally arrayed glomerular capillary domains and efferent, more centrally located domains, with the former constituting most of the capillaries (73,74,75,76,77). There are multiple anastomoses between capillaries within a lobule, and between capillaries in separate lobules, creating a capillary network (Fig. 1.41).
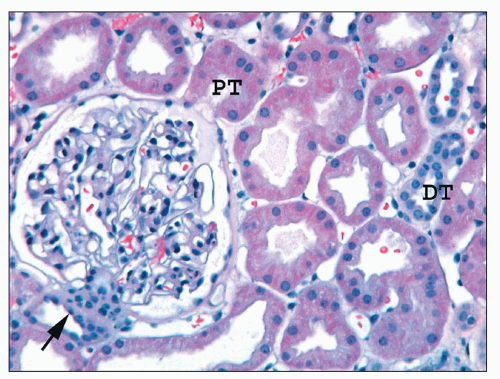 FIGURE 1.39 Human cortex. The macula densa (arrow) and the adjacent extraglomerular lacis cells are on the left; the ostium of the proximal tubule (PT) is on the right. The parietal epithelium lining Bowman capsule is flattened and abruptly transitions to the columnar proximal tubular epithelium at the urinary pole. Most tubules are proximal, but two distal tubules (DTs) are present. (H&E, ×200.) |
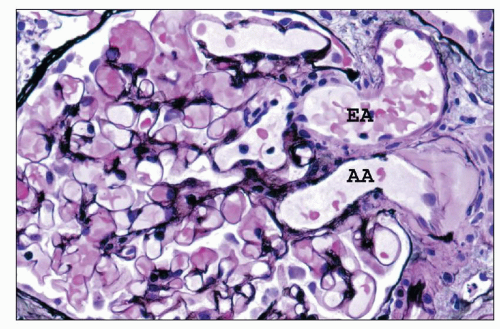 FIGURE 1.40 Human glomerulus with vascular pole. The thin delicate capillary loop basement membranes and slender profiles of mesangial matrix are stained black. The afferent arteriole is identified by the presence of hyalinosis. (Jones methenamine silver, (×400.) AA, afferent arteriole; EA, efferent arteriole. |
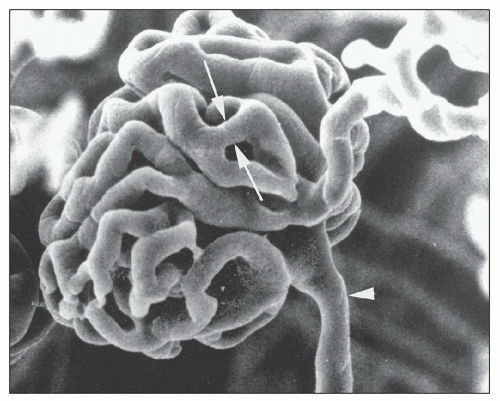 FIGURE 1.41 Scanning electron micrograph of a vascular cast of a rat glomerulus. The afferent arteriole (arrowhead) and efferent arteriole are shown. A capillary loop anastomosis is shown (between arrows). (×450.) (From Gattone VH II, Evan AP, Willis LR, et al. Renal afferent arteriole in the spontaneously hypertensive rat. Hypertension 1983;5:8.) |
of the BC. Parietal cells abruptly transition into columnar proximal tubular cells at the urinary pole (see Figs. 1.38 and 1.39).
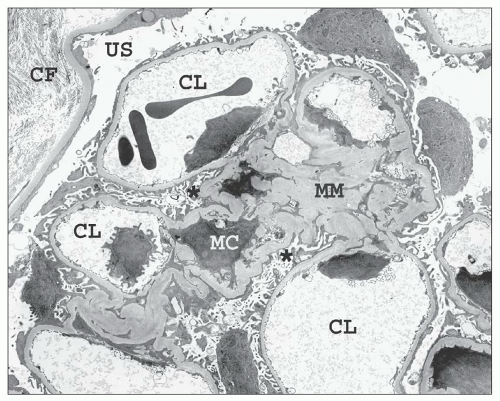 FIGURE 1.42 Transmission electron micrograph of a relatively normal human glomerulus. The mesangial matrix (MM) contains a mesangial cell (MC), and the capillary loops (CLs) are lined by thin fenestrated endothelium. The endothelial cell nuclei are located near the mesangial interface. Podocytes invest both the outer aspects of the capillary loops and the mesangial waist regions (asterisk). The urinary space (US) separates the glomerulus from the Bowman capsule, which is lined by flat parietal epithelial cells. Interstitial collagen fibrils (CF) are visible on the outer aspect of the basal lamina of the Bowman capsule. (×2200.) |
viewing an H&E-stained section under fluorescence (Fig. 1.44). This is useful in kidney biopsies for correlation with immunofluorescence findings (81).
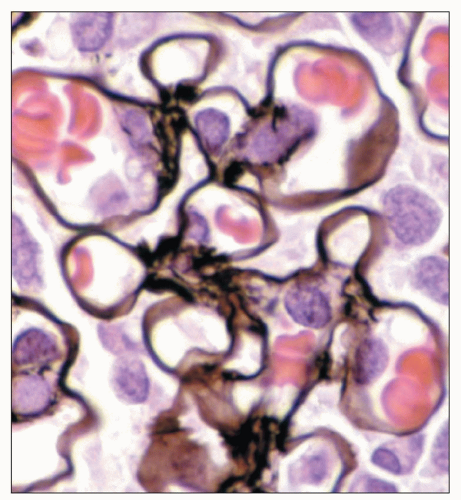 FIGURE 1.43 Normal human glomerulus. The thin delicate detail of the capillary loops and scant mesangium is nicely delineated by the Jones methenamine silver stain. (×650.) |
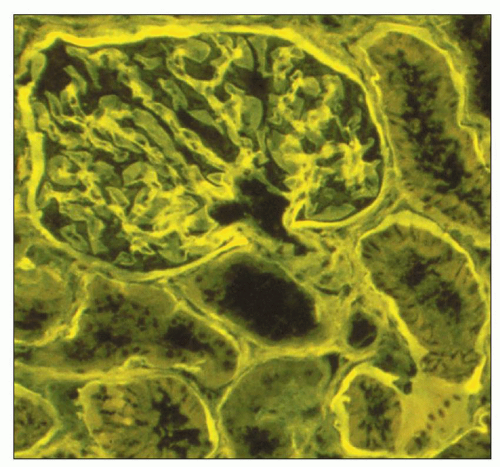 FIGURE 1.44 Normal human cortex. This H&E-stained section is viewed under fluorescent microscopy. The general architectural features of the glomerulus and tubules are easily recognized. (H&E, ×250.) |
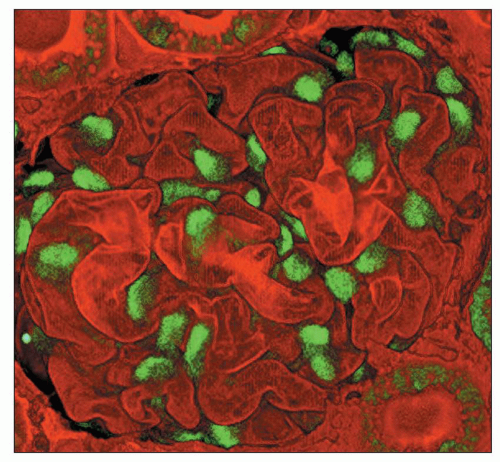 FIGURE 1.45 Two-photon microscopy of a rat glomerulus. Podocytes are the only renal cells of the mature kidney that express WT-1; note the intense nuclear stain. The GBM is stained with Lens culinaris. (×550.) (Photograph courtesy of Carrie Phillips, MD.) |
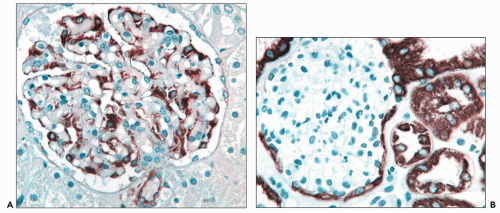 FIGURE 1.46 Human kidney. A: Podocytes express vimentin intermediate filaments while parietal epithelial cells are negative. (Immunoperoxidase stain for vimentin, ×350.) B: Parietal epithelial cells and proximal and distal tubular cells express cytokeratin while podocytes are negative. (Immunoperoxidase stain for cytokeratin 8, ×300.) |
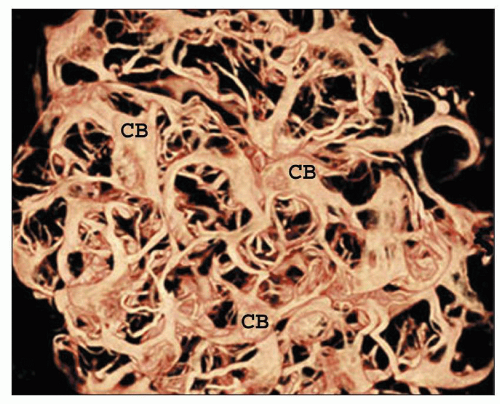 FIGURE 1.47 Two-photon image of a rat glomerulus. Vimentin intermediate filaments are restricted by the cell body (CB) and cell processes. The intervening foot processes are not visible since they lack vimentin. (Immunofluorescent stain for vimentin, ×600.) (Photograph courtesy of Carrie Phillips, MD, and Indiana Center for Biological Microscopy.) |
Related posts:
 Development of the Kidney
Development of the Kidney
 Focal Segmental Glomerulosclerosis
Focal Segmental Glomerulosclerosis
 Renal Involvement in Polyarteritis Nodosa, Kawasaki Disease, Takayasu Arteritis, and Giant Cell Arteritis
Renal Involvement in Polyarteritis Nodosa, Kawasaki Disease, Takayasu Arteritis, and Giant Cell Arteritis
 Pauci-Immune and Antineutrophil Cytoplasmic Autoantibody-Mediated Crescentic Glomerulonephritis and Vasculitis
Pauci-Immune and Antineutrophil Cytoplasmic Autoantibody-Mediated Crescentic Glomerulonephritis and Vasculitis
 Renal Disease Caused by Hypertension
Renal Disease Caused by Hypertension
 Pyelonephritis and Other Direct Renal Infections, Reflux Nephropathy, Hydronephrosis, Hypercalcemia, and Nephrolithiasis
Pyelonephritis and Other Direct Renal Infections, Reflux Nephropathy, Hydronephrosis, Hypercalcemia, and Nephrolithiasis
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


