convenient, should not obviate the need to document the exact renal tract dysmorphology found in a particular patient, nor should it be assumed that there will be one underlying pathogenesis for CAKUT. Thus, the use of the term is analogous to using a disease group designation such as “glomerulonephritis” or “tubulopathy.” As medical and surgical therapies are refined, a new cohort of young babies with severe kidney malformations and renal failure, who might otherwise have perished, are surviving childhood (8,9); as yet, it is too early to know how they will fare through adulthood.
duct. In the female, apart from a few vestigial and inconstant structures, such as the epoophoron, the paroophoron, and Gartner duct, the whole mesonephros degenerates during the 3rd month of gestation.
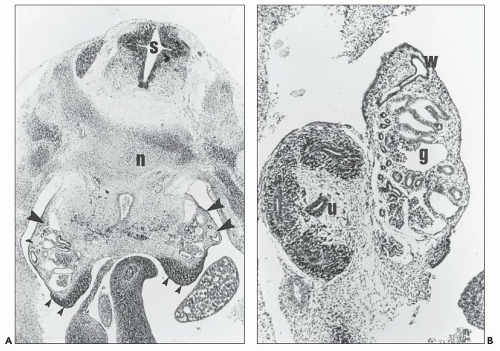 FIGURE 2.1 Early human metanephros and the mesonephros. A: Transverse section of a 5- to 6-week gestation human embryo showing mesonephros (large arrowheads) and relatively undifferentiated gonadal ridge (small arrowheads). Also shown is the spinal cord (s) with the notochord (n) degenerating in a mass of cartilage that will form the vertebral body. (H&E, ×12.5.) B: Six-week human embryo showing the metanephros (left) and the mesonephros (right). The ureteric bud (u) in the center of the metanephros has branched twice, and the mesenchyme is condensing around the branch tips. In contrast, the mesonephros is much more differentiated and contains glomeruli (g) connected to tubules that drain into the wolffian duct (w). (H&E, ×50.) (Courtesy of Dr. P. J. D. Winyard, Institute of Child Health, London, UK.) |
branching is so rapid at this stage, the interstitial portions of the branches are very short, and indeed, sometimes a number of branches appear to arise from a single stem. Distension of the whole system, which is ascribed to the onset of urine production as nephrons start to function, results in the coalescence of the first generations of branches to give the more familiar appearance of the pelvicaliceal system seen postnatally. This is completed by about the 10th to 12th week of gestation. When the minor calyces are formed, some 20 or so ampullae are related to each minor calyx, and it is from these that the papillary collecting ducts originate. Further branching of these ampullae is associated with nephron formation, and these branchings proceed more slowly with longer intervening periods of interstitial growth. From about the 11th week, the spherical expansion of the calyces is limited by the development of nephrons associated with their own and surrounding collecting duct systems that form the developing renal papillae. The minor calyces are indented by the papillae and change from spherical to a wineglass shape, with the cup-shaped portion around the developing papilla and a narrower stem-like infundibular portion connecting to the major calyx. This process is achieved by about the 14th week of gestation
and can be appreciated in histologic sections of the fetal kidneys at this gestational age.
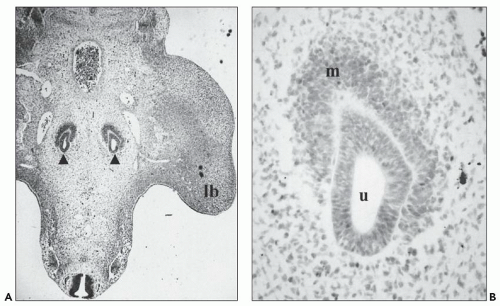 FIGURE 2.2 Human metanephros at the ureteric bud stage. A: Transverse section of a human embryo, approximately 5 weeks’ gestation, showing paired metanephric kidneys (arrowheads), one on each side of the midline. A lower limb bud (lb) is seen on the right (hematoxylin, ×12.5). B: High power of (A) shows one of the ureteric buds (u) capped by the metanephric mesenchyme (m), which is demarcating from the surrounding loosely packed, intermediate mesoderm. |
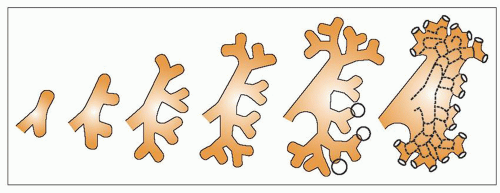 FIGURE 2.3 Development of the renal pelvis. Diagram represents coalescence of the third through fifth generations of branches (circled) of the ureteric bud to form the renal pelvis.) (Modified from Osathanondh V, Potter El. Development of the kidneys as shown by microdissection. II. Renal pelvis, calyces and papillae. Arch Pathol 1963;76:277-289. |
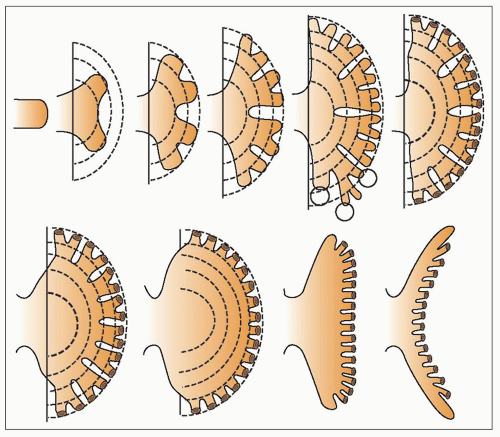 FIGURE 2.4 Development of renal calyces and papillae. Diagrams depict coalescence of the third to fifth generations of branches (circled) of the primordial calyx and inward prolapse of the renal papilla. (Modified from Osathanondh V, Potter El. Development of the kidneys as shown by microdissection. II. Renal pelvis, calyces and papillae. Arch Pathol 1963;76:277-289.) |
arcades. Each ampulla ceases to divide and becomes capable of inducing the formation of a further three to six nephrons. With the induction of further nephrons, the connecting tubule of the older nephron shifts the position of its point of attachment away from the ampulla to the connecting tubule of the next-formed nephron, so they are joined together in a string or arcade of between four and seven nephrons. The innermost and first-formed member of each nephron arcade is a nephron formed during the first period of nephrogenesis and becomes a juxtamedullary nephron in the fully developed organ. The loops of Henle are longest in these nephrons, extending almost to the papillary tip.
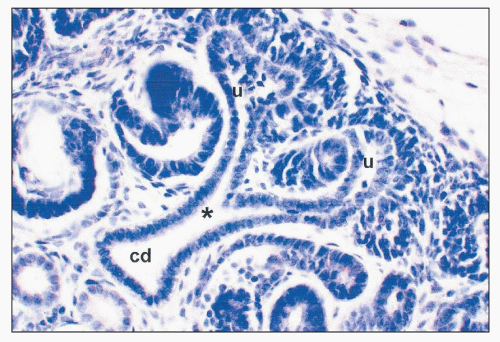 FIGURE 2.5 Metanephros from a 7-week human fetus. A branch point (asterisk) of the ureteric bud/collecting duct (cd) lineage leading to two ureteric bud ampullae (u). (hematoxylin, ×100.) |
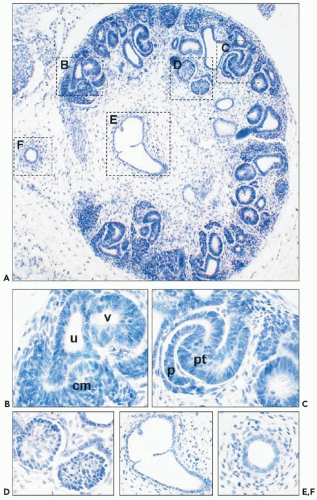 FIGURE 2.6 Metanephros from a 7-week human fetus. A: Low-power view of whole metanephros. Boxed areas are shown in higher powers in subsequent frames. (Hematoxylin, ×12.5.) B: Nephrogenic zone showing the ampulla of a ureteric bud (u) branch, condensed metanephric mesenchyme (cm), and a primitive nephron vesicle (v) that has just undergone the mesenchymal-to-epithelial transition. C: An S-shaped body: the primitive nephron is segmenting into the glomerulus, with cuboidal podocytes (p) with the adjacent proximal tubule (pt). D: The first layer of glomeruli is noted in the deep cortex. E: The renal pelvis lined by a monolayer of the urothelium. F: The ureter consists of a 1-2 cell thick layer of the urothelium surrounded by mesenchyme differentiating into smooth muscle. |
producing the subcapsular so-called nephrogenic zone that can be seen in the fetal kidneys up to 36 weeks and is useful in dating gestational age.
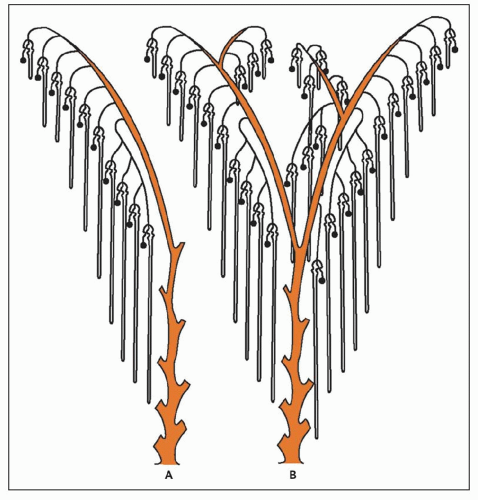 FIGURE 2.7 Arrangement of nephrons at birth as revealed by microdissection. A: Usual pattern with, sequentially, an arcade of four nephrons that drain into a tributary of the collecting duct and six nephrons that drain individually into the collecting duct. B: Possible variations. (Modified from Osathanondh V, Potter El. Development of the kidneys as shown by microdissection. II. Renal pelvis, calyces and papillae. Arch Pathol 1963;76:277-289.) |
that documents the expression patterns of many thousands of gene transcripts through mouse renal tract development (60).
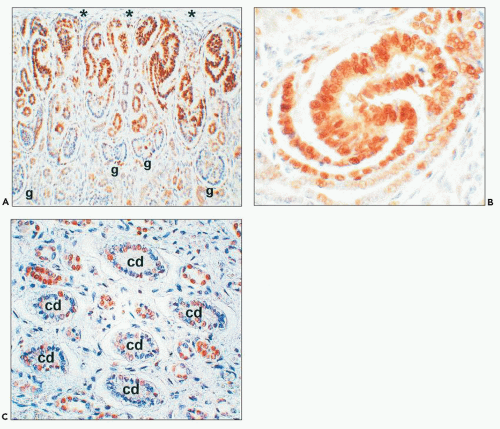 FIGURE 2.8 Cell proliferation in a 25-week human fetal kidney. A: Low-power overview to show that proliferation is prominent in the nephrogenic zone below the renal capsule (asterisks) but is down-regulated deeper in the cortex: Glomeruli are indicated by g. (Section was immunoprobed for proliferating cell nuclear antigen [positive signal appears brown] and counterstained with hematoxylin, ×12.5.) B: Higher power: Most cells within this forming nephron (S-shaped body stage) are proliferating. C: Deep in the same kidney, subsets of nuclei in maturing medullary collecting ducts (cd) are proliferative, as are cells in smaller-caliber tubules, probably representing the loops of Henle, which are growing into the deeper medulla. |
(69), and the PAX family, which contain DNA-binding paired domains (70,71,72) (Fig. 2.9). The specific gene targets of certain of these transcription factors are being investigated. For example, WT1 down-regulates PAX2 (73); up-regulates amphiregulin, a growth factor that stimulates tubule formation in the metanephros (74); and modulates the expression of WNT4, a growth factor implicated in early nephron differentiation (67). Another transcription factor called hepatocyte nuclear factor 1β (HNF1β) promotes the expression of a battery of genes that maintain the differentiated state of renal epithelia; when HNF1β is mutated, renal tubules become cystic (36,65,75,76).
TABLE 2.1 Genetic bases of mouse kidney defects based on mutants | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Related posts:
 Primer on the Pathologic Classification and Diagnosis of Kidney Disease
Primer on the Pathologic Classification and Diagnosis of Kidney Disease
 Focal Segmental Glomerulosclerosis
Focal Segmental Glomerulosclerosis
 Renal Involvement in Polyarteritis Nodosa, Kawasaki Disease, Takayasu Arteritis, and Giant Cell Arteritis
Renal Involvement in Polyarteritis Nodosa, Kawasaki Disease, Takayasu Arteritis, and Giant Cell Arteritis
 Pauci-Immune and Antineutrophil Cytoplasmic Autoantibody-Mediated Crescentic Glomerulonephritis and Vasculitis
Pauci-Immune and Antineutrophil Cytoplasmic Autoantibody-Mediated Crescentic Glomerulonephritis and Vasculitis
 Renal Disease Caused by Hypertension
Renal Disease Caused by Hypertension
 Pyelonephritis and Other Direct Renal Infections, Reflux Nephropathy, Hydronephrosis, Hypercalcemia, and Nephrolithiasis
Pyelonephritis and Other Direct Renal Infections, Reflux Nephropathy, Hydronephrosis, Hypercalcemia, and Nephrolithiasis
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


