of two different families, the src (including p59fyn and p56lck) and ZAP 70 families.2 The CD45 protein, a tyrosine phosphatase, contributes to the activation process by dephosphorylating an autoinhibitory site on the p56lck PTK. Intracellular domains of several TCR/CD3 proteins contain activation motifs that are crucial for antigen-stimulated signaling. Certain tyrosine residues within these motifs serve as targets for the catalytic activity of src family PTKs. Subsequently, these phosphorylated tyrosines serve as docking stations for the SH2 domains (recognition structures for select phosphotyrosinecontaining motifs) of the ZAP-70 PTK. Following antigenic engagement of the TCR/CD3 complex, select serine residues of the TCR and CD3 chains are also phosphorylated.2,5
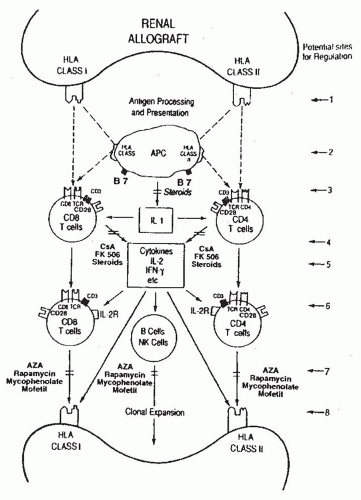 FIGURE 81.1 The antiallograft response. Schematic representation of human leukocyte antigens (HLA), the primary stimuli for the initiation of the antiallograft response, cell-surface proteins participating in antigenic recognition and signal transduction, contribution of the cytokines and multiple cell types to the immune response,and the potential sites for the regulation of the antiallograft response.Site /:Minimizing histocompatibility between the recipients and the donor (e.g., HLA matching).Site 2: Prevention of monokine production by antigen-presenting cells (e.g., corticosteroids). Site 3: Blockade of antigen recognition (e.g.,OKT3 mAbs).S/fe4: Inhibition of T cell cytokine production (e.g., cyclosporin A [CsA]).Sife 5: Inhibition of cytokine activity (e.g.,anti-interleukin-2 [IL-2] antibody). S/fe 6: Inhibition of cell cycle progression (e.g.,anti-IL-2 receptor antibody). Site 7: Inhibition of clonal expansion (e.g.,azathioprine [AZA]). Site 8: Prevention of allograft damage by masking target antigen molecules (e.g., antibodies directed at adhesion molecules). HLA class I: HLA-A, B, and C antigens; HLA class II: HLA-DR, DP,and DQ antigens. IFN-γ, interferon-y; NK cells, natural killer cells. |
[NF-AT], nuclear factor kappa B [FN-κB] , activator protein 1 [AP-1]) and the transcriptional activation and expression of genes central to T cell growth (e.g., interleukin-2 [IL-2] and receptors for IL-2 and IL-15).2,5,12
TABLE 81.1 Cellular Elements Contributing to the Antiallograft Response | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||
powerful T cell costimulus.19 A subset of monocytes and dendritic cells constitutively express CD80 and CD86 at low levels and cytokines (e.g., granulocyte-macrophage colony-stimulating factor [GMCSF] or interferon-γ [IFN-γ]) stimulate heightened expression of CD80 and CD86 on monocytes, B cells, and dendritic cells.19 Many T cells express B7-binding proteins (i.e., CD28 proteins that are constitutively expressed on the surface of CD4 + T cells and CTLA-4 [CD152]), a protein whose ectodomain is closely related to that of CD28, and is expressed upon activated CD44- and CD84- T cells. CD28 binding of B7 molecules stimulates a Ca2+-independent activation pathway that leads to stable transcription of the IL-2, IL-2 receptors, and other activation genes resulting in vigorous T cell proliferation.19 For some time, the terms CD28 and the costimulatory receptor were considered synonymous by some, but the demonstration that robust T cell activation occurs in CD28-deficient mice indicated that other receptor ligand systems contribute to signal two.20 In particular, the interaction between CD40 expressed upon APCs and CD40 ligand (CD154) expressed by antigen-activated CD44- T cells has received great attention as a potent second signal.21
TABLE 81.2 Cell-Surface Proteins Important for T Cell Activationa | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
of APCs then represent target molecules for the regulation of the antiallograft response. Indeed, transplantation tolerance has been induced in experimental models by targeting a variety of cell-surface molecules that contribute to the generation of costimulatory signals, and tolerance to histoincompatible human kidney allografts has been accomplished with a conditioning regimen that includes monoclonal antibodies directed at the CD2 protein.23
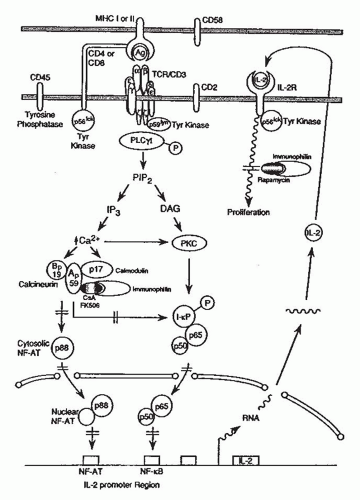 FIGURE 81.2 Signal transduction in T cells and mechanisms of action of cyclosporin A (CsA), FK-506, or rapamycin.Signaling molecules and transmembrane signaling events participating in the transduction of antigenic signals from the plasma membrane of the T cells to the nucleus are schematically shown. The sites of action of the drug (CsA/FK-506/rapamycin)-immunophilin complex are also shown.kg, antigen; Ap59 and Bp19, subunits of calcineurin;DAG, diacylglycerol; l-κB, inhibitory factor kappa B;IL.-2, interleukin-2; immunophilin, cyclophilin or FK-binding protein; IP3, inositol 1,4,5-triphosphate; MHC, major histocompatibility complex; NF-AT, nuclear factor of activated T cells; NF-κB, nuclear factor kappa B; P, phosphotyrosine; PIP2, phosphatidylinositol 4,5-biphosphate; PKC, protein kinase C;PLCγ1, phospholipase C gamma-1; Tyr kinase, tyrosine kinase. (Adapted from Schreier MH, Baumann G, Zenke G, et al. Inhibition of T-cell signaling pathways by immunophilin drug complexes: Are side effects inherent to immunosuppressive properties? Transplant Proc 1993;25:502.) |
PTK activity. The ligand-activated common γ chain recruits a member of the Janus kinase family, Jak 3, to the receptor complex that leads to activation of a member of the STAT family Activation of this particular Jak-STAT pathway is essential for the proliferation of antigen-activated T cells. The subsequent events leading to IL-2/IL-15-dependent proliferation are not fully resolved; however, IL-2/IL-15—stimulated expression of several DNA binding proteins including Bcl-2, c-jun, c-fos, and c-myc contributes to cell cycle progression.31,32 It is interesting and probably significant that IL-2, but not IL-15, triggers apoptosis of many antigen-activation T cells. In this way, IL-15-triggered events may be more detrimental to the antiallograft response than those initiated by IL-2. As IL-15 is not produced by T cells, IL-15 expression is not regulated by cyclosporine or tacrolimus.
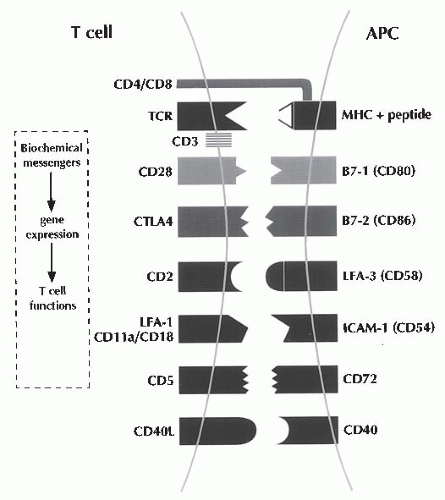 FIGURE 81.3 T cell/antigen-presenting cell contact sites. In this schema of T cell activation, the antigenic signal is initiated by the physical interaction between the clonally variant T cell antigen receptor (TCR) α-β-heterodime, and the antigenic peptide displayed by MHC on antigen-presenting cells (APCs).The antigenic signal is transduced into the cell by the CD3 proteins.The CD4 and the CD8 antigens function as associative recognition structures, and restrict TCR recognition to class II and class I antigens of MHC, respectively. Additional T cell- surface receptors generate the obligatory costimulatory signals by interacting with their counterreceptors expressed on the surface of the APCs.The simultaneous delivery to the T cells of the antigenic signal and the costimulatory signal results in the optimum generation of second messengers (such as calcium), expression of transcription factors (such as nuclear factor of activated T cells), and T cell growth-promoting genes (such as IL-2).The CD28 antigen as well as the CTLA4 antigen can interact with both the B7-1 and B7-2 antigens.The CD28 antigen generates a stimulatory signal, and CTLA4, unlike CD28, generates a negative signal. CD, cluster designation; ICAM-1, intercellular adhesion molecule-1; LFA-1, leukocyte function-associated antigen-1;MHC, major histocompatibility complex. (From Suthanthiran M.Transplantation tolerance: fooling mother nature. Proc Natl Acad Sci U S A. 1996;93:12072.) |
and antibody producing B cells/plasma cells (Fig. 81.1). Cytokines facilitate not only the T cell effector arm and TCR but also the B cell/plasma cell arm by promoting the production of cytopathic antibodies. Moreover, cytokines such as IFN-y and tumor necrosis factor-α (TNF-α) can amplify the ongoing immune response by upregulating the expression of HLA molecules as well as costimulatory molecules (e.g., B7) on graft parenchymal cells and APCs (Fig. 81.1). We and others have demonstrated the presence of antigen-specific cytotoxic T lymphocytes (CTL) and anti-HLA antibodies during or preceding a clinical rejection episode.35,36 We have detected messenger RNA (mRNA) encoding the CTL-selective serine protease (granzyme B), perforin, Fasligand attack molecules, and immunoregulatory cytokines, such as IL-10 and IL-15, in human renal allografts undergoing acute rejection.37 Indeed, these gene expression events may anticipate clinically apparent rejection. More recent efforts to develop a noninvasive method for the molecular diagnosis of rejection have proved rewarding. Using either peripheral blood38 or urinary leukocytes39 rejection-related, gene expression events evident in renal biopsy specimens are robustly detected in peripheral blood or urinary sediment specimens. Initial results from large-scale multicenter trials (e.g., Clinical Trials in Organ Transplantation, CTOT-04) support the hypothesis that noninvasive diagnosis of acute TMR is feasible by measurement of genes encoding cytotoxic attack molecules in urine, and the urinary cell mRNA profiles may anticipate the future development of acute TMR.40 We speculate as well that a noninvasive, molecular diagnostic approach to rejection would be of value toward the detection of insidious, clinically silent rejection episodes that, although rarely detected through standard measures, are steroid-sensitive but usually lead to chronic rejection.41
TABLE 81.3 Mechanisms of Action of Small Molecule Immunosuppressantsa | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||
were comparable (4.5% vs. 3.8%) after a 6-month observation period in the MMF (n = 22) or AZA (n = 26) arm. The trial participants received cyclosporine and prednisone as maintenance immunosuppressive therapy and antithymocyte globulin induction was used in 27% of the recipients maintained on MMF and 46% in the AZA conversion group. It is worth noting that high-risk patients including retransplant recipients, highly sensitized, and those with a history of steroid-resistant rejection were all excluded from the trial.50
Related posts:
 Computed Tomography and Magnetic Resonance Imaging
Computed Tomography and Magnetic Resonance Imaging
 Alport Syndrome, Fabry Disease, and Nail-Patella Syndrome
Alport Syndrome, Fabry Disease, and Nail-Patella Syndrome
 Pathophysiology of Nephrotoxic Cell Injury
Pathophysiology of Nephrotoxic Cell Injury
 Immunoglobulin A Nephropathy and Henoch-Schönlein Purpura
Immunoglobulin A Nephropathy and Henoch-Schönlein Purpura
 Monoclonal Gammopathies: Multiple Myeloma, Amyloidosis, and Related Disorders
Monoclonal Gammopathies: Multiple Myeloma, Amyloidosis, and Related Disorders
 Peritoneal Dialysis
Peritoneal Dialysis
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


