Chapter 9 Liver immunology
Fundamentals of Immunology
The immune system is composed of two major components: innate and adaptive immunity (Fig. 9.1; Janeway, 2001). Innate immune cells are armed with hard-wired detection systems that recognize common structures found on pathogens or changes in surface molecules of host cells that signal danger. Such pattern recognition receptors (PRRs) account for the rapid response of innate immune cells to infection or host injury. A variety of PRRs exist, of which the best characterized is the Toll-like receptor (TLR) family. In humans, 11 TLRs are currently known that recognize various structurally conserved components of bacteria and viruses in addition to endogenous intracellular ligands. Engagement of PRRs by their respective ligands causes activation of innate immune cells, such as neutrophils, monocytes, macrophages, natural killer (NK) cells, and dendritic cells (DCs). The ensuing response results in destruction of the invading pathogen or tumor via phagocytosis or release of various cytotoxic or inflammatory agents.
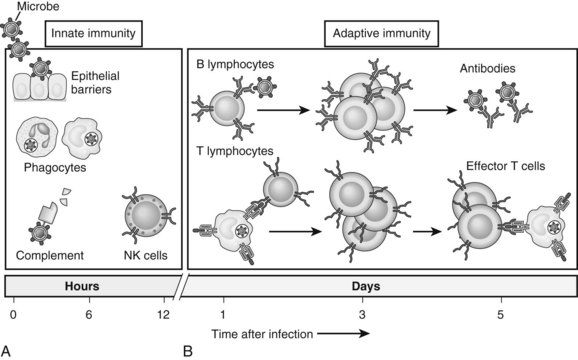
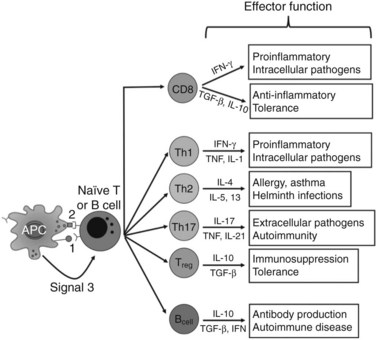
In contrast, adaptive immunity refers to the arm of the immune system that is involved in antigen-specific responses, which occur either later in a particular immune response or rapidly after repeat exposure to a particular pathogen. The adaptive immune system is essentially comprised of T and B cells that circulate within the blood, lymphatic tissues, and organs. These lymphocytes interact with various antigen-presenting cells (APCs) of the innate immune system, such as DCs and macrophages. Activation occurs in the presence of specific signals between APCs and T or B cells. These signals include 1) stable antigen presentation within the context of major histocompatibility complex (MHC) molecules MHC-I or MHC-II, 2) appropriate costimulation by APCs and the corresponding receptor on T or B cells, and 3) cytokine-mediated signals that can modulate the overall response (Fig. 9.2). Innate and adaptive immune responses provide protection against a vast array of pathogens, preserving normal cells and tissues from attack. Failure of the immune system’s ability to correctly recognize foreign or abnormal cells may facilitate the development or spread of infection or malignancy. In contrast, inability to distinguish self from foreign antigens may result in autoimmune disease.
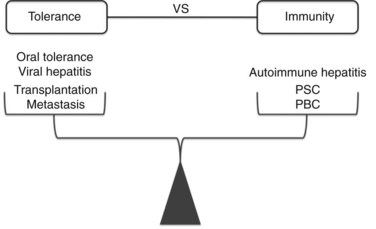
Modern clinical and experimental work has shown that the liver is particularly important in immunity (Fig. 9.3). First, the fact that the liver is one of the most common sites for metastatic disease suggests that it may have an increased propensity toward immunologic acceptance. Second, allogeneic liver transplantation can be accomplished in certain strains of mice without the need for immunosuppression, and in humans it often requires less immunosuppression compared with kidney or other solid organ transplants. Third, the liver is home to chronic viral infection in the form of hepatitis B and C. Additionally, oral ingestion or portal vein injection of foreign proteins can lead to tolerance in animal models. Conversely, the liver is the site of several autoimmune processes, including primary sclerosing cholangitis and primary biliary sclerosis. Despite the significant role that the liver plays in altering the balance between tolerance and immunity, the study of liver immunology remains in its infancy. This chapter discusses our current understanding of the function of liver immune cells and their role in disease.
Anatomic Considerations
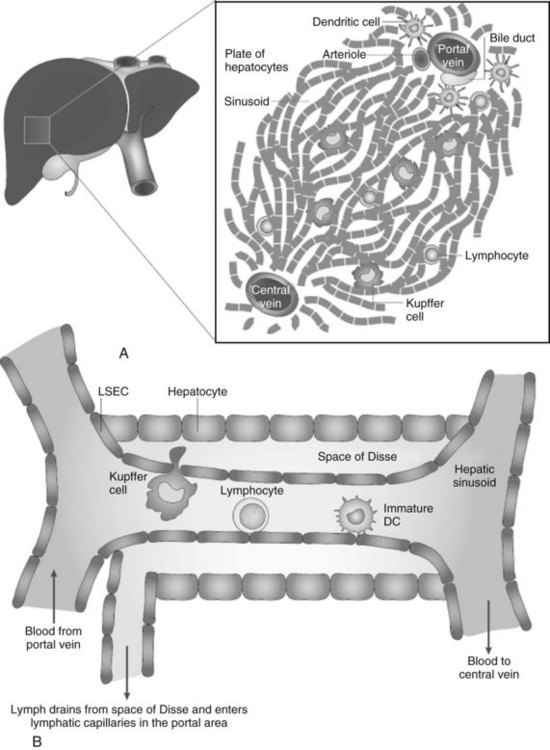
Because the vascular supply of the liver derives principally from the portal venous system draining the gut, a heavy antigen load is delivered to the liver, particularly after meals. Portal venous blood flows slowly through the vast network of hepatic sinusoids, which are discontinuously lined by fenestrated endothelium lacking a basement membrane (Fig. 9.4; see also Chapter 6). The sluggish flow of blood allows for the efficient capture of antigens by leukocytes traveling in the blood within the sinusoids and by the endothelial cells lining the sinusoids. The microscopic anatomy of the liver also favors the ability of blood-borne leukocytes to interact with hepatic parenchymal cells and resident immune cells of the liver.
Tolerance Versus Immunity
One of the most intriguing aspects of liver immunology is the propensity of antigens passing through the liver to produce tolerance rather than immunity. In experimental animal models and clinical studies of humans undergoing liver transplantation, a greater propensity for graft acceptance has been noted compared with transplantation of other solid organs. A liver transplant protects a kidney allograft transplanted simultaneously from the same donor (Creput et al, 2003). Trying to decipher the reasons for the liver’s inherent propensity for tolerance has been a particularly active area of research in the field of transplantation and cancer immunobiology.
Multiple competing theories to explain the cause of liver tolerance have been advanced. One postulate is that, as in central thymic tolerance, clonal deletion of antigen-specific T cells occurs in the liver. Another theory is that liver allografts release large amounts of soluble MHC class I molecules that potentially could function to neutralize donor-specific antibodies or cytolytic T cells. Kupffer cells (KCs) and natural killer (NK) T cells have been implicated as being important in the development of hepatic tolerance, because the depletion of either has been associated with a loss of oral tolerance. Liver DCs also have been suggested to play a role in liver tolerance, because as a group they have been shown to be less immunogenic than DCs from lymphoid organs such as the spleen (Bamboat et al, 2009). In addition, the numbers of several unusual cell types are elevated in the liver, and more recent studies have shown that multiple lineages of intrahepatic leukocytes are unique in function and phenotype compared with their counterparts from lymphoid organs. Controversy exists, however, as to whether these differences are due to de novo development, conditioning within the liver, or chemotactic signals in the liver that attract a unique population of cells. Regardless, the differences in composition and function are likely to play an important role in defining the nature of liver immunology. We devote a portion of this chapter to the major populations of liver immune cells and their role in various liver diseases.
Liver Immune Cells
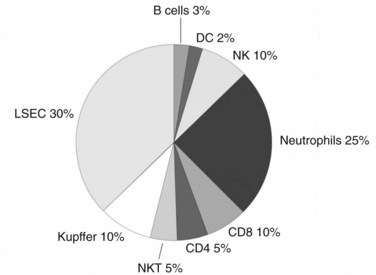
Although the liver is primarily a nonlymphoid organ, it contains numerous nonparenchymal cells (NPCs; see Chapter 6), one quarter of which are leukocytes. The composition of the intrahepatic leukocyte population is markedly different from that seen in other organs (Fig. 9.5). As in the rest of the body, however, the liver contains most of the cellular components of innate and adaptive immunity. Advances in technology have allowed for the precise delineation and isolation of cell types based on the expression of a distinct set of surface markers.
Antigen-Presenting Cells
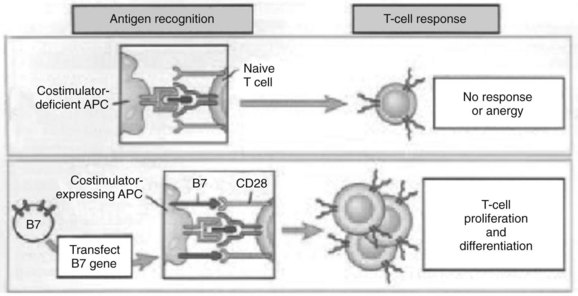
The experimental observation that antigens passing through the liver can lead to tolerance makes the understanding of antigen presentation in the liver particularly germane (Li et al, 2004). Antigen-presenting cells (APCs) play a crucial role in determining the nature of adaptive immune responses. The manner in which an APC presents any particular antigen can alter dramatically the response of antigen-specific T cells. Specifically, when antigen presentation occurs in conjunction with the appropriate costimulatory molecules, T cells proliferate and develop an immunogenic phenotypic and functional profile (Fig. 9.6). In contrast, antigens presented in the absence of costimulation lead to anergy or activation-induced T-cell death, two of the mechanisms of peripheral tolerance induction and maintenance.
Dendritic Cells
The body of literature studying the function of DCs grown in vitro from murine bone marrow progenitors or human peripheral blood mononuclear cells is vast. DCs isolated directly from the spleen, lymph nodes, or thymus also have been well studied. In contrast, because of the rarity of DCs in the liver, few studies have shown the phenotype or function of liver DCs. Initial studies focused on DC progenitors grown from liver nonparenchymal cells cultured with various cytokines, including granulocyte-macrophage colony-stimulating factor (GM-CSF). These early studies showed that liver DC progenitors are relatively immature cells with poor immunostimulatory ability that can promote immunologic tolerance and prolong solid organ transplants in animal models (Lu et al, 1994, 2001; Lau & Thomson, 2003).
To gain a better understanding of liver DCs as they function in situ, recent focus has been on studying freshly isolated liver DCs. In our initial studies, we wished to increase the number of liver DCs to facilitate their isolation. Because GM-CSF had been used to grow DC progenitors in culture from liver nonparenchymal cells, and because we had previously shown that in vivo GM-CSF overexpression increases the number of myeloid DCs in the spleen (Miller et al, 2002a), we suspected that it might expand liver DCs as well. We found that overexpression of GM-CSF using an adenoviral vector led to a dramatic increase in the number of highly immunostimulatory liver DCs (Pillarisetty et al, 2003). We also showed that a large proportion of the cells recruited to the liver by GM-CSF were DC precursor cells that develop into CD11c+ DCs in culture, which suggests that DC development can occur in the liver. We used immunohistochemistry to locate both types of recruited cells. The liver DCs were concentrated around the central veins, whereas the DC precursors were distributed diffusely throughout the liver parenchyma.
Our experience with expanding liver DCs sheds light on in situ liver DC development, but we wished to understand the phenotype and function of liver DCs in normal, untreated mice. When our ability to isolate liver DCs from normal mice improved, we began to study them in depth. We found that compared with the relatively well-studied DCs from the spleen, CD11c+ liver DCs were immature and only weakly immunostimulatory (Pillarisetty et al, 2004). We also noticed that, in contrast to spleen DCs, liver DCs are heterogeneous in their expression of MHC class II and costimulatory molecules. We further separated CD11c+ liver DCs based on their expression of the myeloid marker CD11b and the lymphoid marker CD8α+, which have been commonly used to define murine DC subtypes. Myeloid (CD11b+) and lymphoid (CD8α+) liver DCs, which each comprise approximately 10% of the total population of DCs in the liver, were as able to activate T cells as were their splenic counterparts. The bulk of the remaining cells, which had low to no expression of CD11b and CD8α, were only poorly immunostimulatory. This study showed that the presence of these atypical DCs was primarily responsible for the weakly activating nature of liver DCs on the whole. More recently, we discovered that despite having multiple APCs, the CD11chi subset of DCs within the liver are required for effective presentation of soluble protein (Plitas et al, 2008). Using a transgenic mouse in which CD11chi DCs can be depleted selectively, we found that activation of antigen-specific CD8+ T cells in the liver only occurred in the presence of CD11chi DCs.
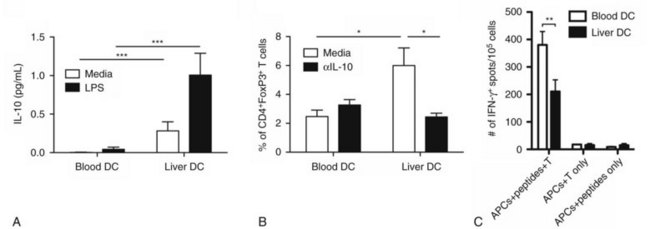
Our investigation into liver DCs has now been extended into humans. We found that freshly isolated DCs from human liver exhibit tolerogenic properties when compared with autologous blood DCs. Liver DCs are weaker stimulators of T cells and favor the production of antiinflammatory IL-10, which induces the differentiation of naïve CD4+ T cells into regulatory T cells with suppressive function (Fig. 9.7; Bamboat et al, 2009a).
Kupffer Cells
Liver macrophages, referred to as Kupffer cells (KCs), have long been believed to be the primary phagocytic cells of the liver. KCs represent the largest pool of macrophages in the body, derived from monocytic precursors in the blood. They are typically found in the hepatic sinusoids; however, they also can migrate through the space of Disse to interact with hepatocytes (see Fig. 9.4). KCs have been thought to play a major role in antigen presentation and have been implicated in portal venous tolerance. Evidence also suggests that KCs may regulate T-cell responses to antigens and induce immune tolerance in the setting of liver allografts (Sun et al, 2003).
Liver Sinusoidal Endothelial Cells
Several studies have shown that in addition to serving as the building blocks lining the sinusoids, LSECs are immune cells with the ability to capture and present antigen and activate T cells (Knolle & Limmer, 2001). As with KCs, considerable controversy surrounds the immunologic function of LSECs. In contrast to earlier work, we have shown more recently that although LSECs are highly capable of capturing various antigens in vivo and in vitro, they lack the ability to activate T cells in the absence of exogenous costimulation (Katz et al, 2004). The differences in results may derive from the use of more specific methods of cell isolation in the latter study. The finding that LSECs are not independently capable of triggering a T cell–mediated immune response does not, however, exclude the possibility that LSECs, in concert with DCs or KCs, play an important role in antigen presentation in the liver.
Effector Cells
T Cells
Liver T cells are divided into the same types of cells, as are the cells of lymphoid organs, however, evidence shows differences in the proportions of T-cell subtypes found within the liver. The interaction between CD4+ and CD8+ cells is clearly important, and more recent studies have suggested that the ratio between these two cell types—which is approximately 2 : 1 in the spleen, lymph nodes, and peripheral blood—is reversed in the liver (Crispe, 2003). Preliminary murine data from our laboratory, using more inclusive isolation techniques, have shown that the liver has a similar CD4+ to CD8+ T-cell ratio of approximately 1.5 : 1 as found in the spleen and node (Katz et al, 2005). Prior studies have shown that unconventional T cells are highly represented in the liver compared with the spleen or lymph nodes, and our data are in agreement with these findings.
γδ T cells
The γδ T-cell receptor is relatively invariant, but it can recognize multiple nonpeptide antigens without the need for MHC presentation. γδ T cells represent 10% of liver T cells, whereas they comprise only a small proportion (< 5%) of T cells in the blood or lymphoid organs. γδ T cells also are highly represented in the skin and at mucosal interfaces and are thought to play a regulatory role through their secretion of activating and modulatory cytokines as part of an early response to atypical bacterial and viral pathogens. They can influence the activation of γδ T cells and also have been shown to be crucial in tumor immunity through their early secretion of interferon γ (IFN-γ) (Gao et al, 2003). The high proportion of γδ T cells in the liver suggests that they have an important immunologic role, but further investigation is required. Similarly, both human and mouse livers are enriched with a large proportion of memory T cells. Although some investigators believe this is because the liver serves as a reservoir for activated T cells just before they die, other theories include that the liver secretes chemokines that attract CD8+ T cells.
Natural Killer T Cells
Related posts:
 Preoperative and postoperative nutrition in hepatobiliary surgery
Preoperative and postoperative nutrition in hepatobiliary surgery
 Portal hypertension in children
Portal hypertension in children
 Liver transplantation for cholangiocarcinoma and other neoplastic diseases
Liver transplantation for cholangiocarcinoma and other neoplastic diseases
 Distal splenorenal shunt
Distal splenorenal shunt
 Magnetic resonance imaging of the liver, biliary tract, and pancreas
Magnetic resonance imaging of the liver, biliary tract, and pancreas
 Interventional endoscopy: Technical aspects
Interventional endoscopy: Technical aspects
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


