individual molecule is random, a concentration gradient is required for any net transfer of molecules to occur across a membrane. Thus, the concentration gradient represents the driving force for net transport.
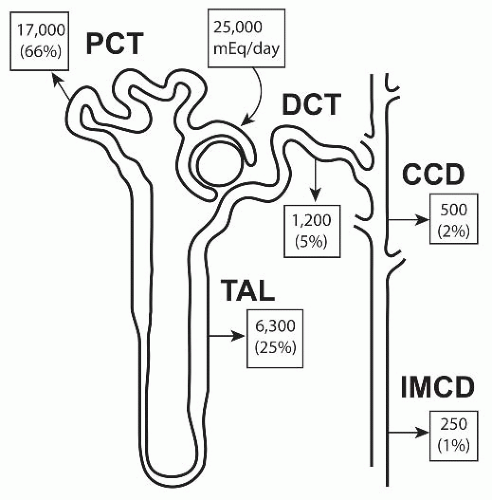 FIGURE 5.1 The contribution of various nephron segments to Na+ transport. PCT, proximal convoluted tubule; DCT, distal convoluted tubule; CCD, cortical collecting duct; TAL, thick ascending limb; IMCD, inner medullary collecting duct. |
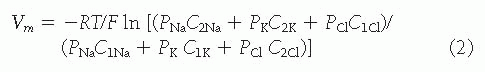
TABLE 5.1 Inherited Disorders of Renal Sodium Transport | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
segment approach zero.26,27 The omission of glucose and amino acids from luminal fluids reduces both the potential difference and the volume absorptive rate.24,28 In the S3 segment, on the other hand, the omission of glucose and alanine has no effect either on the potential difference or on the fluid absorptive rate,25,29 although the deletion of all organic solutes does reduce volume absorption by 50%.30 The rates of transport of glucose, amino acids, phosphate, and Na+ in the early proximal convoluted tubule exceed those in the proximal straight tubule.31,32,33 These rates correlate well with the relative basolateral membrane areas of the respective segments.34
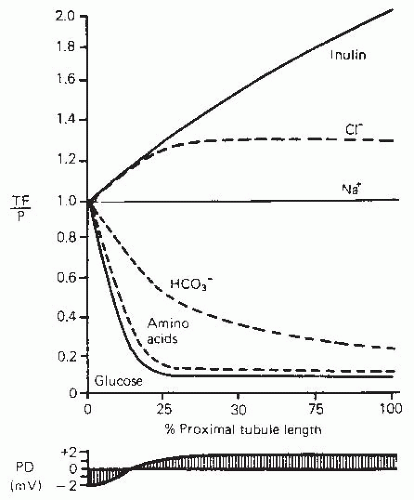 FIGURE 5.2 The profile of transepithelial voltage and solute concentrations along the mammalian proximal tubule. TF/P, tubular fluid/plasma concentration ratio; PD, potential difference. (From: Rector FC Jr. Sodium, bicarbonate, and chloride absorption by the proximal tubule. Am J Physiol. 1983;244:F461, with permission.) |
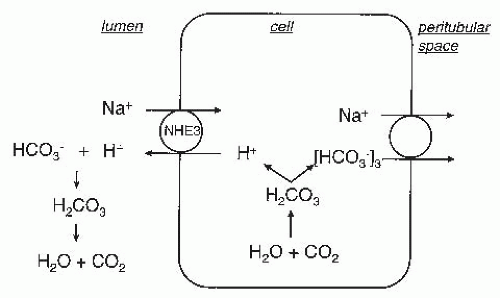 FIGURE 5.3 The scheme of NaHCO3 transport mediated by Na+/H+ exchange. See text for explanation. |
three separate transport systems,73 one that transports all neutral amino acids, one specific for imino acids, and one for the β-amino acids. Glycine may also have a specific transporter.74 In the kidney, neutral amino acid transport is driven by a Na+ gradient, as supported by experiments in slices, perfused tubules, and brush border membranes. The neutral amino acid transporter B0AT1 (SLC6A19) cotransports one Na+ per amino acid.75 The Km of the substrate decreases with an increasing cosubstrate concentration and vice versa. The initial step for transport involves the binding of the amino acid to B0AT1, and this binding affinity increases under hyperpolarizing conditions.76
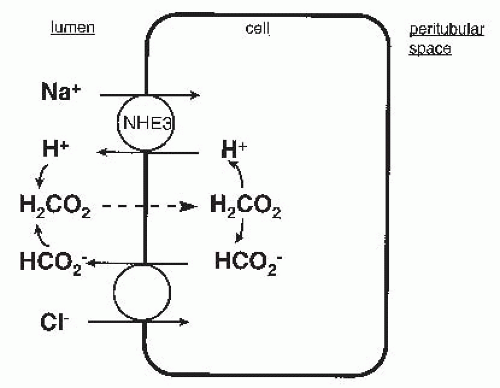 FIGURE 5.4 The scheme of neutral NaCl transport mediated by the parallel action of Na+-H+ exchange and formate/Cl– exchange. Formate (HCO2–) combines with H+ in the tubular lumen to form formic acid (H2CO2), which reenters the cell by nonionic diffusion. A similar scheme applies for oxalate-Cl– exchange. |
and HCO3– that enter the cell are thought to be recycled through the [Na+(HCO–3)3]2- cotransporter (see previous). Indeed, Na+(HCO–3)2/Cl– exchange may account for much more Cl– movement than KCl transport.108 Na+-independent Cl–/HCO3– exchange is also present in the basement membrane of proximal tubules.106,107,108 However, under physiologic conditions, this process mediates net Cl– influx and does not contribute to net NaCl absorption.
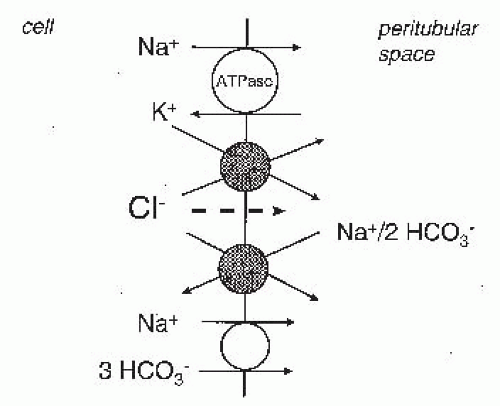 FIGURE 5.5 The transport pathways for Na+ and Cl– absorption across the basolateral membrane of proximal tubular cells. Cl– can leave the cell via KCl cotransport, Na+-2HCO3–/Cl– exchange, and Cl– channels (minor). Na+ exits via the Na+,K+-ATPase and Na+-3(HCO3–) cotransport. |
does not inhibit the exchange activity of NHE3.132 The current model for this inhibition is that NHE3 associates with PKA indirectly via NHERF and the cytoskeletal protein ezrin. PKA, when active, phosphorylates NHE3 at serines 552 and 605, which mediates the inhibition of the exchanger.133 NHE3 is directly phosphorylated by other protein kinases, including calmodulin-dependent protein kinase II, which inhibits Na+/H+ exchange activity and PKC, which stimulates the exchanger.134
force to account for the observed rates of fluid reabsorption.20 Experimental evidence supports the view that absolute luminal hypotonicity is a significant driving force for fluid reabsorption in the proximal tubule. Thus, when proximal tubules are perfused and bathed by symmetric NaCl solutions, the luminal fluid becomes slightly hypotonic.155 The development of luminal hypotonicity can be amplified by maneuvers that decrease the water permeability of the proximal tubule. The aquaporin 1 (AQP1) water channel is abundantly expressed in the proximal tubule. In AQP1 knockout mice, the osmolality of tubular fluid at the end of the proximal tubule is significantly lower than in normal mice.156 As the luminal fluid becomes more hypotonic, the resorbate becomes more hypertonic, and the degree of resorbate hypertonicity correlates with the rate of volume reabsorption by the tubules.157
ionic conductance and low water permeability effectively further dilutes fluid entering the TAL from the ascending thin limb.
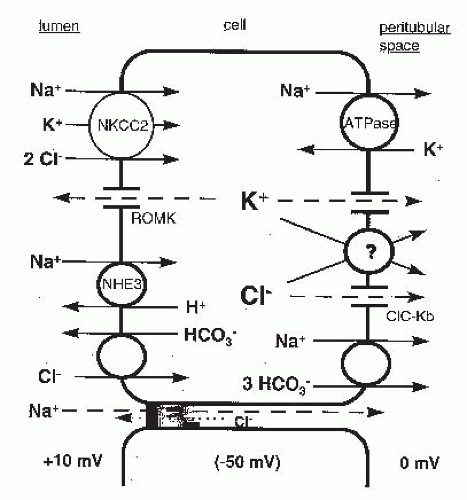 FIGURE 5.6 A model depicting the major elements of the mechanism of NaCl absorption by the thick ascending limb. Dashed lines indicate passive ion movements down electrochemical gradients. ROMK, renal outer medullary K+ channel; ClC-Kb, chloride channel Kb. |
chain reaction (RT-PCR) studies demonstrated the expression of NKCC2 in the MTAL and CTAL,195 and immunohistochemical studies indicate that NKCC2 is localized to the apical membrane of these nephron segments.196 The importance of NKCC2 in mediating salt reabsorption in the TAL is illustrated by the fact that loss-of-function mutations of NKCC2 cause Bartter syndrome (Table 5.1),197 a Mendelian salt-wasting disorder characterized by hypokalemia, metabolic alkalosis, hyperaldosteronism, and normal-to-low blood pressure, results from a defect in salt absorption by the thick ascending limb.
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


