Chapter 1 The Hypothalamic-Pituitary-Ovarian Axis and Control of the Menstrual Cycle
INTRODUCTION
The hypothalamus and pituitary gland form a unit that exerts control over a wide range of endocrine organs, including the gonads. This chapter describes the hypothalamic-pituitary-ovarian axis and control of the menstrual cycle, which is modulated by the central nervous system, other endocrine systems, and the environment. Key hormones in the hypothalamic-pituitary-ovarian axis include gonadotropin-releasing hormone (GnRH), follicle-stimulating hormone (FSH), luteinizing hormone (LH), estradiol, and progesterone (Table 1-1). Supporting roles are also played by inhibin, activin, follistatin, and endorphins.
THE HYPOTHALAMUS
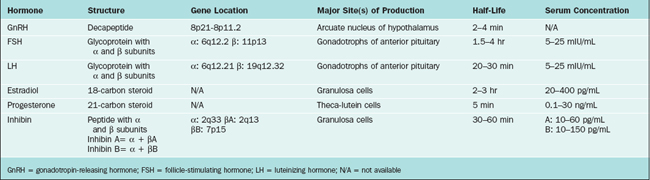
The hypothalamus forms the lower part of the lateral wall and the floor of the third ventricle, and weighs approximately 10 grams. The hypothalamus is typically divided into eight specific nuclei (consistently clustered groups of neurons) and three areas (less clustered, less distinctly demarcated neurons), as illustrated in Figure 1-1. From a reproductive standpoint, the most important of these are the arcuate nucleus and the preoptic area, the principal sites of GnRH-producing neurons.1 The arcuate nucleus is located in the medial basal hypothalamus and is the most proximal of all the hypothalamic nuclei to the optic chiasm and the pituitary stalk. The arcuate nucleus is also the site of dopamine-secreting neurons that function to inhibit pituitary prolactin secretion and neurons that secrete growth hormone-releasing hormone.
GnRH
GnRH is the primary hypothalamic regulator of pituitary reproductive function. Two human forms of GnRH (GnRH-I and GnRH-II) have been identified.2,3 Both are decapeptides and are the products of different genes. At least 20 other types of GnRH have been identified in fish, amphibians and protochordates, but none of these are believed to be present in humans.4,5
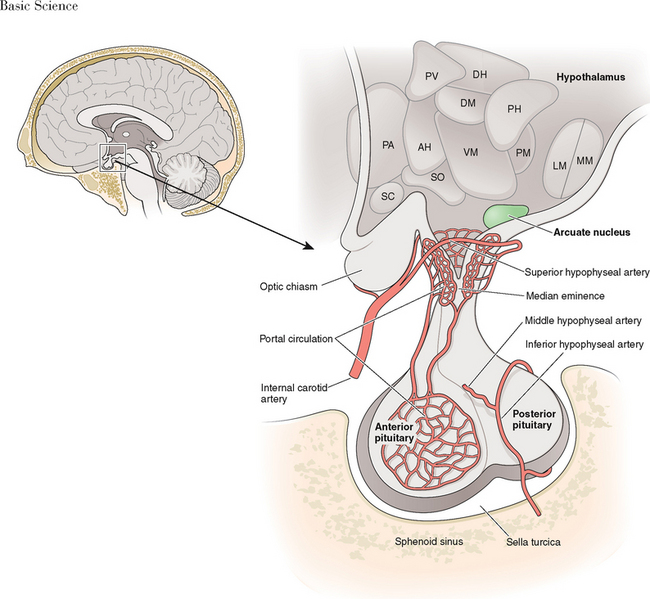
GnRH-I was first characterized and synthesized in 1971 by Andrew Schally and Roger Guillemin, an accomplishment for which both men ultimately received the Nobel prize.6–9 The structure of GnRH-I is common to all mammals, and its action is similar in males and females (Fig. 1-2). GnRH-I is synthesized from a much larger, 92-amino acid precursor peptide that contains GnRH-associated peptide.10 GnRH-I then travels along an axonal pathway called the tuberoinfundibular tract to the median eminence of the hypothalamus, where it is released into the portal circulation in a pulsatile fashion. The half-life of GnRH-I is very short (2 to 4 minutes) because it is rapidly cleaved between amino acids 5 and 6, 6 and 7, and 9 and 10. Because of its short half-life and rapid dilution in the peripheral circulation, serum GnRH-I levels are difficult to measure and do not correlate with pituitary action.
GnRH-I has three principal actions on anterior pituitary gonadotrophs: (1) synthesis and storage of gonadotropins, (2) movement of gonadotropins from the reserve pool to a point where they can be readily released, and (3) direct secretion of gonadotropins. GnRH-I pulses occur in response to intrinsic rhythmic activity within GnRH neurons in the arcuate nucleus. Pulsatile release of GnRH-I from the median eminence within a critical frequency and amplitude results in normal gonadotropin secretion.11,12 Continuous, rather than pulsatile, exposure to GnRH-I results in suppression of FSH and LH secretion and suppression of gonadotropin gene transcription.13,14
In the absence of gonadal feedback, the GnRH pulse frequency is approximately once per hour.15 During the menstrual cycle the frequency and amplitude of GnRH pulses vary in response to hypothalamic feedback (Table 1-2).16 In general, the follicular phase is characterized by high-frequency, low-amplitude pulses, and the luteal phase is characterized by lower-frequency, higher-amplitude pulses.17,18 However, considerable variability exists both between and within individuals.19 In humans, GnRH-I pulse frequency and amplitude are best approximated by measuring LH pulse frequency and amplitude.
Table 1-2 Menstrual Cycle Variation in LH Pulse Frequency and Amplitude
| Cycle Phase | Mean Frequency (minutes) | Mean Amplitude (mIU/mL) |
|---|---|---|
| Early follicular | 90 | 6.5 |
| Mid-follicular | 50 | 5 |
| Late follicular | 60–70 | 7 |
| Early luteal | 100 | 15 |
| Mid-luteal | 150 | 12 |
| Late luteal | 200 | 8 |
GnRH-II is most highly expressed outside of the brain, in tissues that include kidneys, bone marrow, and prostate. This is in contrast to GnRH-I, which is not expressed in high levels outside the brain. Although GnRH-II can induce release of both FSH and LH, it appears to have a wide array of physiologic functions outside the brain including regulation of cellular proliferation and mediation of ovarian and placental hormone secretion.20
Initial attempts in the mid-1990s to identify estrogen receptors in GnRH neurons were unsuccessful.21,22 However, subsequent use of more sophisticated techniques identified estrogen receptors α and β in the arcuate nucleus.23–26 Both receptors mediate estrogen action on GnRH neurons in vivo.27,28 The GnRH gene contains a hormone response element for the estrogen–estrogen receptor complex.29 Transcription of GnRH-I and GnRH-II is differentially regulated by estrogen.30 The regulatory role of estradiol on GnRH is complex. Estrogen inhibits GnRH gene expression/biosynthesis, but secretion of GnRH may be increased, decreased, or unaffected.31,32
The activity of the hypothalamus is further modulated by nervous stimuli from higher brain centers. GnRH neurons exhibit many connections to each other and to other neurons. Some of the neurotransmitters that modulate GnRH secretion are outlined in Table 1-3. The effects of these neurotransmitters help explain the mechanism by which certain physical or clinical conditions may affect the menstrual cycle (Table 1-4).
Table 1-3 Neurotransmitter Effects on GnRH Release
| Neurotransmitter | Effect |
|---|---|
| Dopamine | Inhibits GnRH release |
| Endorphin | Inhibits GnRH release |
| Serotonin | Inhibits GnRH release |
| Norepinephrine, epinephrine | Stimulates GnRH release |
Table 1-4 Mechanisms for Oligo/amenorrhea in Various Clinical Conditions
| Hyperprolactinemia | Elevated dopamine suppresses GnRH |
| Hypothyroidism | Elevated TRH increases prolactin, which in turn increases dopamine which, then suppresses GnRH |
| Stress | Increased corticotropin (ACTH) results in increased endorphins (both are derived from the same peptide precursor); endorphins suppress GnRH |
| Exercise | Increased endorphins suppress GnRH |
TRH = thyrotropin-releasing hormone; GnRH = gonadotropin-releasing hormone.
Cells that produce GnRH originate embryologically from the olfactory area.33 GnRH neurons, like olfactory epithelial cells of the nasal cavity, have cilia.34 During embryogenesis GnRH neurons migrate from the medial olfactory placode to the arcuate nucleus of the hypothalamus.35 The common origin of GnRH and olfactory neurons is demonstrated by Kallmann’s syndrome, where GnRH deficiency is associated with anosmia. Kallmann’s syndrome is believed to be caused by a variety of gene defects that affect neuronal cell migration.36
The common origin of GnRH and olfactory neurons is also suggestive of the relationship between pheromones and menstrual cyclicity. Pheromones are small airborne chemicals that when secreted externally by one individual may be perceived by other individuals of the same species, producing a change in sexual or social behavior. It is well recognized that women who work or live together often develop synchrony of their menstrual cycles.37 Moreover, it has been shown that odorless compounds from the axillae of cycling women can alter the cycle characteristics of recipient women.38 Presumably, these alterations occur through olfactory GnRH-mediated mechanisms.
The GnRH Receptor
The GnRH-I receptor is a G-protein receptor that utilizes inositol triphosphate and diacylglycerol as second messengers to stimulate protein kinase, release of calcium ions, and cyclic adenosine monophosphate (cAMP) activity. The GnRH-I receptor is encoded by a gene on chromosome 14q21.1 and is expressed in many parts of the body outside of the brain, including ovarian follicles and the placenta. In humans it appears that GnRH-II signaling occurs through the GnRH-I receptor.5 Although a GnRH-II receptor is present in many mammalian species, its functional capacity is limited due to a frame shift and a stop codon. GnRH receptors are regulated by many substances, including GnRH itself, inhibin, activin, estrogen, and progesterone.
Changes to the amino acid sequence of GnRH can extend its half-life to hours or days and can change its biologic activity from an agonist to an antagonist. All of the GnRH agonists currently available extend their half-life by substitutions at amino acid 6 and sometimes amino acid 10 of native GnRH (Table 1-5). Continuous activation of the GnRH receptor results in desensitization due to phosphorylation and conformational change of the receptor, uncoupling from G proteins, internalization of the receptor via endocytosis, and decreased receptor synthesis.39,40 On administration all GnRH agonists increase gonadotropin secretion (the flare effect). However, after 7 to 14 days GnRH receptor desensitization occurs and pituitary suppression is achieved.
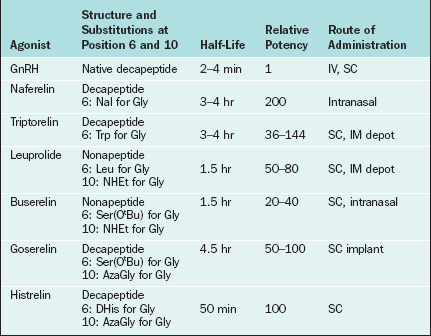
In contrast, GnRH antagonists directly inhibit gonadotropin secretion. Structurally, GnRH antagonists are characterized by multiple amino acid substitutions to the natural GnRH decapeptide. The commercially available GnRH antagonists cetrorelix and ganirelix have large amino acid additions to position 1 of native GnRH. GnRH antagonists compete for and occupy pituitary GnRH receptors, thus competitively blocking endogenous GnRH–GnRH receptor binding. In contrast to GnRH agonists, there is no flare effect with GnRH antagonists. Because receptor loss does not occur, a constant supply of antagonist is necessary to ensure that all GnRH receptors are continuously occupied. Thus, the therapeutic dosage range for antagonists is typically higher than that for agonists (mg versus μg).
THE PITUITARY
The pituitary gland measures approximately 15 × 10 × 6 mm and weighs approximately 500 to 900 mg. The pituitary gland lies immediately beneath the third ventricle and just above the sphenoidal sinus in a bony cavity called the sella turcica (Turkish saddle). It consists of anterior and posterior lobes, each having different embryologic origins, functions, and control mechanisms (Fig. 1-3).
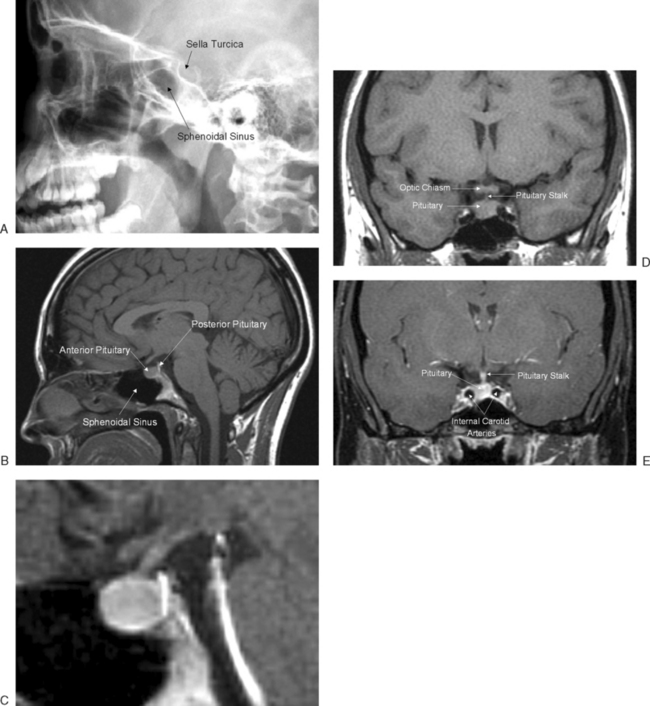
The Anterior Pituitary
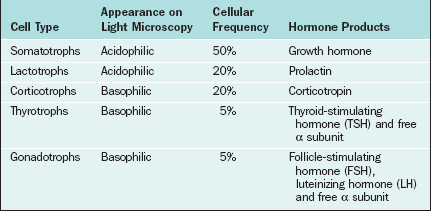
The major cell types of the anterior pituitary are outlined in Table 1-6. The anterior pituitary is a classic endocrine gland in that it is composed of secretory cells of epithelial origin supported by connective tissue rich in blood and lymphatic capillaries. In accordance with their active synthetic function the endocrine cells are characterized by prominent nuclei and prolific mitochondria, endoplasmic reticulum, Golgi bodies, and secretory vesicles. Synthesis of gonadotropins takes place in the rough endoplasmic reticulum, after which the hormones are packaged within the Golgi apparatus and stored as secretory granules. In response to GnRH the secretory granules are extruded from the cell membrane. The endothelial lining of capillary sinusoids is fenestrated, facilitating the passage of pituitary hormones into the sinusoids.
Gonadotropins
FSH
The sialic acid content of FSH, LH, TSH, and hCG varies, and these differences are largely responsible for variations in half-life of the glycoprotein hormones. The liver is the major site of clearance for gonadotropins. Sialic acid prevents hepatic clearance; thus, the greater the sialic acid content, the longer the half-life.41 hCG, with 20 sialic acid residues, has the longest half-life (about 24 hours), whereas LH (1 to 2 sialic acid residues) has the shortest half-life (20 to 30 minutes). Addition of sialic acid residues in urinary-derived commercially available gonadotropins (e.g., hMG) is responsible for their longer half-life (30 hours).
In gonadotrophs of the anterior pituitary, GnRH signaling leads to transcription of the α and β subunits for both FSH and LH. The GnRH-dependent availability of the β subunits is the rate-limiting step in gonadotropin synthesis. Although both FSH and LH require GnRH stimulation, synthesis of the FSH β subunit also requires the presence of activin.42,43
FSH and LH Receptors
In contrast, FSH receptors are found exclusively on granulosa cells in the ovary. FSH binds to receptors on the cell surface of granulosa cells in antral follicles. Like LH, FSH acts via the cAMP-dependent protein kinase pathway.44 In response to FSH the androgens produced as a result of LH stimulation are then aromatized to estrogens in granulosa cells.
Opioid Modulation of Pituitary Hormones
Opioids (i.e., endogenous opiates) are natural occurring sedative narcotics produced in the brain whose structure and function are similar to opium. Opioids include enkephalins, endorphins, and dynorphins; they modulate every pituitary hormone by acting on the hypothalamus. An important action of opioids is to inhibit gonadotropin secretion by suppressing GnRH release.45
Opioid tone is an important regulator of menstrual cyclicity.46–49 Endorphins are at a nadir in the early follicular phase (menstruation) and gradually rise to peak levels in the luteal phase in response to the rise in estrogen and progesterone. It is believed that opioids mediate the negative feedback of ovarian steroids on gonadotropin release, particularly in the luteal phase.50
Endogenous opioids appear to play a central role in hypothalamic amenorrhea. Treatment of women suffering from this condition with an opioid receptor antagonist (e.g., naltrexone) results in the return to ovulatory menstrual patterns and even conception in some cases.51,52 Women with stress-related amenorrhea demonstrate increased hypothalamic corticotropin-releasing hormone and pituitary corticotropin, which manifests as hypercortisolism.53 The corticotropin precursor peptide, pro-opiomelanocortin, is also the precursor for endorphin synthesis. It is hypothesized that stress-related amenorrhea is the result of GnRH inhibition secondary to increased production of endogenous opioids. Opioids also rise during exercise (“runners’ high”), and this may contribute to hypothalamic amenorrhea in athletes.54,55
Ovarian Peptide Hormone Feedback on Gonadotropin Secretion
The ovary secrets two polypeptide hormones that inhibit or stimulate FSH secretion by the anterior pituitary. Inhibin and activin act as opposing nonsteroidal gonadal hormones that regulate FSH synthesis and secretion by the pituitary. They also have paracrine effects within the ovary, where they modulate follicle growth and steroidogenesis. Follistatin is a binding protein that modulates the effects of activin but not inhibin.
Inhibin and Activin
Inhibin and activin are members of the transforming growth factor-β (TGF-β) superfamily of ligands, which includes müllerian inhibiting substance (MIS; see Chapter 2). Like the gonadotropins, inhibin and activin are comprised of two subunits. Inhibin is comprised of an α and β subunit and has been isolated in two forms containing different β subunits, Inhibin-A and Inhibin-B. Activin is comprised of two beta subunits identical to those found in inhibin.
Inhibin is secreted by granulosa cells in response to FSH.56 However, mRNA for inhibin has also been found in pituitary gonadotrophs. Inhibin selectively inhibits FSH but not LH secretion.57 Thus, a negative feedback loop is created where FSH stimulates inhibin and in turn inhibin suppresses FSH.
Inhibin B is predominantly secreted in the follicular phase of the menstrual cycle, whereas inhibin A is predominantly secreted in the luteal phase.58 Peak levels of inhibin B in the follicular phase are in the range of 50 to 100 pg/mL. Peak levels of inhibin A in the luteal phase are between 40 and 60 pg/mL.
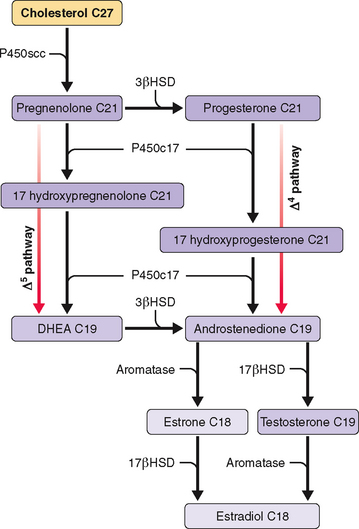
OVARIAN STEROIDOGENESIS DURING THE MENSTRUAL CYCLE
Ovarian steroidogenesis during the menstrual cycle occurs in granulosa and theca cells (Table 1-7 and Fig. 1-4). Before ovulation, theca cells are separated from granulosa cells in the same follicle by a basal membrane. Thus, granulosa cells of preovulatory follicles do not have a blood supply. However, at the time of the LH surge the preovulatory follicle undergoes luteinization with disappearance of the basal membrane and capillary invasion of the granulosa cells. Theca cells become theca-lutein cells, and granulosa cells become granulosa-lutein cells.
Table 1-7 Site of Synthesis of Major Steroidogenic Products of the Ovary
| Cell Type | Major Steroid Hormone Products |
|---|---|
| Theca cells | Androgens (androstenedione, DHEA, testosterone)* |
| Granulosa cells | Estrogens (estradiol, estrone) |
| Theca-lutein cells | Progestogens (progesterone, 17-hydroxyprogesterone)** |
| Granulosa-lutein cells | Estrogens (estradiol, estrone) |
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


