Chapter 15 Polycystic Ovary Syndrome
INTRODUCTION
In 1935, Stein and Leventhal described patients who had a constellation of amenorrhea, infertility, and enlarged, polycystic ovaries.1 They found that bilateral ovarian wedge resection resulted in regular menses and even pregnancy in some. They reasoned that the thickened tunica was preventing follicles from reaching the surface of the ovary, resulting in the classic appearance of an enlarged ovary with multiple follicles beneath the cortex. Today we know that the mere finding of a polycystic-appearing ovary is not pathognomonic of PCOS, because many females without the disorder will have similar ovarian morphology.2,3
DIAGNOSTIC CRITERIA
In 1990, a conference on PCOS sponsored by the National Institutes of Health (NIH) led to diagnostic criteria based on a majority opinion of conference participants.4 The criteria included: hyperandrogenism and/or hyperandrogenemia, chronic anovulation, and exclusion of other known disorders. In 2003, the European Society of Human Reproduction and Embryology (ESHRE)/American Society of Reproductive Medicine (ASRM)-sponsored PCOS consensus workshop in Rotterdam revised the diagnostic criteria for PCOS.5 The revised criteria state that PCOS remains a diagnosis of exclusion, but that two out of the following three criteria must be present: (1) oligo-ovulation or anovulation, (2) hyperandrogenism and/or hyperandrogenemia, and (3) polycystic ovaries.
PREVALENCE
The prevalence of PCOS is estimated to be 4% to 12% of reproductive-age women. The largest U.S. study on PCOS prevalence was published in 1998.6 Out of 277 women included in the study, 4.0% had PCOS as defined by the 1990 NIH criteria. The prevalence was 4.7% for white women and 3.4% for black women. The inclusion of polycystic ovaries in the 2003 Rotterdam criteria calls for re-evaluation of the prevalence of PCOS, because 21% to 23% of normal women have polycystic-appearing ovaries on ultrasound.2,3
CLINICAL PRESENTATION
Hyperandrogenism
Clinical manifestations of hyperandrogenemia include hirsutism, acne, and male pattern alopecia. Hirsutism is defined as the growth of coarse, pigmented hairs in androgen-dependent areas such as the face, chest, back, and lower abdomen. Approximately 80% of hirsute patients will have PCOS.7 The modified Ferriman-Gallway scoring system can be used for clinical assessment of hirsutism. This was originally used in the United Kingdom for a population of presumably white women, and scores hair growth in nine body areas from 0 (absence of terminal hairs) to 4 (extensive terminal hair growth).8 Sex hair growth differs among various ethnic and genetic groups, being decreased in Asians, even when high androgen levels are present.9
Other hyperandrogenic manifestations commonly found in PCOS patients include acne and alopecia.10,11 Acne is a result of androgen stimulation of the pilosebaceous unit with increased skin oiliness.10 Cela and colleagues investigated a multiethnic group of 89 women who presented with androgenic alopecia and found that 67% had polycystic ovaries, compared to 27% of controls.11 In addition, women with alopecia had higher androgen levels and a higher prevalence of hirsutism.
Obesity
Obesity is very common in PCOS, with the android pattern present in approximately 44% of women with PCOS.12 This central obesity is more characteristic of PCOS, because these patients have an increased waist-to-hip ratio compared to obese women without PCOS.13 Hyperinsulinemia may stimulate central adiposity, which, in turn, exacerbates underlying or latent insulin resistance.14 It has been shown that obese women with PCOS have greater insulin resistance than weight-matched controls.15
Acanthosis Nigricans
Acanthosis nigricans is a dermatologic condition of hyperkeratosis and increased skin pigmentation with raised, symmetric, darkened, velvety plaques that commonly appear on the nape of the neck. It can also be found in the axilla, groin, and other intertriginous areas of the body. This skin finding is a common manifestation of insulin resistance in patients with PCOS. Mor and coworkers found that acanthosis nigricans is more likely to be found in PCOS patients with insulin resistance compared to PCOS patients without insulin resistance (OR = 6.0, P < 0.5).16 The elevated insulin level has a mitogenic effect on basal cells of the epidermis, making acanthosis nigricans a relatively specific clinical marker of insulin resistance.17 Other pathologic conditions associated with acanthosis nigricans include insulinoma and malignant disease, especially adenocarcinoma of the stomach.
Reproductive Aberration
Miscarriage
The risk of a first-trimester spontaneous abortion (i.e., miscarriage) is reported to be significantly higher for patients with PCOS than in normal women. The spontaneous abortion rate in PCOS is reported to be 30%.18 In comparison, retrospective studies find the risk of spontaneous abortion to be 5% to 14% for normal women.19,20 Of patients with recurrent miscarriage, 36% to 82% have polycystic ovaries.18,21,22
Several explanations have been offered. For example, Homburg and colleagues demonstrated that high concentrations of LH during the follicular phase in women with polycystic ovaries have a deleterious effect on rates of conception and are associated with early pregnancy loss.23 In another study, those with elevated LH levels had a 65% miscarriage rate compared to 12% of pregnancies with normal LH levels.24 Elevated serum androgen levels, obesity, and hyperinsulinemia have also been implicated as risk factors for early pregnancy loss.25,26
Polycystic Ovarian Morphology
The Rotterdam ESHRE/ASRM-sponsored PCOS consensus workshop group included the polycystic morphology of the ovary as one of the diagnostic criteria for the syndrome.5 A polycystic ovary is defined as having 12 or more follicles in one ovary measuring 2 to 9 mm in diameter, and/or increased ovarian volume of greater than 10 mL, which is the maximum size of a normal ovary3 (Fig. 15-1).
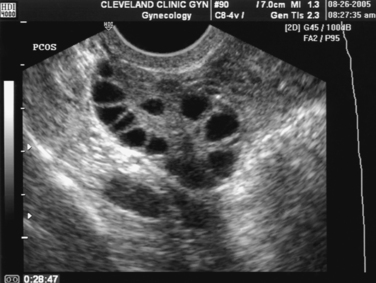
This definition should not be substituted with a subjective appearance of a polycystic ovary. The classic image is that of an enlarged ovary containing an increased number of small follicles around the periphery of the cortex (i.e., a string of pearls) with a bright echogenic stroma. The characteristic increase in stromal volume differentiates the polycystic ovary from a multifollicular ovary.3
The follicle distribution is omitted from the definition of a polycystic ovary, as well as the increase in stromal echogenicity and volume, but ovarian volume has been shown to be a good surrogate for stromal volume.27 Oral contraceptives modify ovarian morphology; thus the definition of a polycystic ovary does not apply to women taking these medications. When there is a dominant follicle (>10 mm) or a corpus luteum, the ultrasound should be repeated during the early follicular phase of the next cycle.
PATHOGENESIS
Altered Gonadotropin Secretion
One of the well-described features of PCOS is an increase in LH and relative decrease in follicle stimulating hormone (FSH).28 The relative decrease in FSH is the chief cause of anovulation, because increasing FSH will lead to folliculogenesis. The pulsatile secretion of LH from the pituitary is increased in amplitude and frequency in PCOS.29 In addition, the pituitary has a greater LH response to gonadotropin-releasing hormone (GnRH) compared with normal women.29,30 Elevations in LH could be secondary to increased GnRH pulse frequency, stimulatory factors other than GnRH, or a combination of these effects.
The pulsatile secretion of GnRH cannot be studied in humans, so it must be inferred by detecting peripheral LH patterns. A study of PCOS women by Berga and colleagues found increased pulse frequency and amplitude for LH and α-subunit, providing evidence for aberrant increases in GnRH pulse frequency (Fig. 15-2).29 Furthermore, increased GnRH pulse frequency in rats has been shown to increase LHβ gene expression.31 Elevated LH is not caused by altered pituitary sensitivity to GnRH; GnRH receptor blockade resulted in similar LH decreases in PCOS and normal women.32
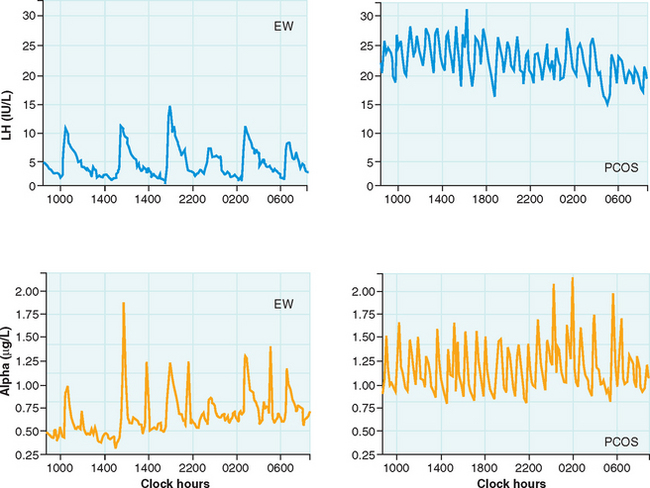
These findings suggest a derangement of the hypothalamic-pituitary axis, which appears to play a major role; many of the cardinal features of PCOS can be traced to alterations in gonadotropins. The basis for decreased FSH secretion in PCOS has not been determined, although the negative feedback effect of chronic unopposed estrogen secretion in these women has been implicated as a mechanism.33
Altered GnRH Drive
Neuroanatomic Considerations
The GnRH pulse generator refers to the synchronized pulsatile secretion of GnRH from neurons that are widely distributed in the medial basal hypothalamus. GnRH is synthesized in neuronal cell bodies that have migrated during fetal life from the olfactory placode to the hypothalamus and is secreted from neuroendocrine terminals localized in the median eminence. Knobil conducted experiments with the Rhesus monkey to establish that the GnRH system exhibits rhythmic electrical behavior in the arcuate nucleus of the medial basal hypothalamus.34 There was remarkable synchrony between pulses of GnRH in the portal blood and LH pulses in peripheral blood. This phenomenon was later studied in isolated human medial basal hypothalamus where GnRH pulses were found to occur at a frequency of 60 to 100 minutes.35 Thus, the GnRH neurons per se, with their intrinsic pulsatile secretion, appear to constitute the GnRH pulse generator.
There is compelling evidence that non-neuronal cells, such as glial and endothelial cells, regulate GnRH secretion in the median eminence. Glial processes belonging to either astrocytes or tanycytes separate GnRH nerve endings from the pericapillary space in the median eminence and, through signaling molecules such as nitric oxide, play a role in regulation of GnRH secretion into pituitary portal blood vessels.36,37 The secretion of GnRH into the portal vasculature also appears to be regulated by dynamic remodeling of GnRH neurovascular junctions. Morphologic plasticity of the median eminence during the menstrual cycle has been demonstrated, where the maximal number of GnRH neurovasculature junctions are found during the LH surge.38
In Vitro GnRH Neuroregulation
Mellon and coworkers developed a cell line of immortalized hypothalamic GnRH neurons (GT1 cells) that were determined to have an inherent ability to secrete GnRH in a pulsatile fashion.40 The development of GnRH cell lines, such as the GT1 cells, has permitted numerous studies to identify putative factors that regulate pulsatile GnRH secretion. Substances implicated in the modulation of the GnRH pulse generator include norepinephrine, dopamine, insulin-like growth factor I (IGF-I), γ-aminobutyric acid (GABA), and opioids, among others. There also appears to be an autocrine regulation of GnRH release, because GT1 cells themselves express GnRH receptors. The activation of GnRH receptors enhances action potentials, heightens synchronization of neuronal activities, and results in pulsatile release of GnRH.
In Vivo GnRH Neuroregulation in PCOS
The GnRH pulse generator in PCOS patients is intrinsically faster, and the frequency is less likely to be suppressed with continuous estrogen and progesterone treatment.40 A persistently rapid GnRH pulse generator would increase LH secretion and could suppress FSH levels low enough to prevent folliculogenesis.
Increased central adrenergic tone has been implicated as a cause of the aberrations of GnRH and gonadotropin secretion in PCOS. One possible mechanism is the increase in local blood flow and permeability of the portal vascular system, permitting the entry of increased amounts of GnRH.41 Dopamine injection into the third ventricle led to a rapid increase in GnRH and prolactin inhibitory factor in portal blood, suggesting dopamine-mediated regulation of GnRH and prolactin inhibitory factor.42 The identification of β1-adrenergic and D1-dopaminergic receptors on GT1 GnRH neurons provides a mechanism by which norepinephrine and dopamine could regulate gonadotropin release via direct synapses on GnRH neurons.43
Other studies have examined the effect of dopamine on immunoreactive LH. Dopamine exerts an inhibitory effect on LH secretion in normally cycling women44 and also has been shown to lower circulating LH in PCOS patients.45 It has been suggested that an impairment of the dopaminergic inhibitory activity on LH secretion leads to the increased frequency of LH pulses seen in PCOS women.46 In fact, the dopamine agonist bromocriptine has been shown to lower circulating LH and improve menstrual function in normoprolactinemic PCOS women.47 This finding has not been consistently demonstrated, as Lobo and coworkers did not find an increase in the ratio of bioactive to immunoreactive LH in response to dopamine alone or a dopamine-carbidopa combination, and concluded that dopamine did not play a critical role in LH secretion.48
The role of IGF-I in modulation of GnRH cells has also been investigated. IGF-I regulates growth, differentiation, survival, and reproductive function. The IGF receptor is a tyrosine kinase receptor located in the periphery and central nervous system, including the median eminence.49 Mouse studies have shown that IGF-I mRNA in the hypothalamus increases through peripubertal and adult development.50 IGF-I was found to stimulate GnRH gene expression in postnatal and peripubertal mice.50 Another study using the GT1 cell line demonstrated that IGF-I treatment caused a significant increase in GnRH nuclear primary transcript levels and cytoplasmic mRNA.51 In PCOS women, an increased ratio of IGF-I to IGF binding proteins correlated significantly with increased concentrations of circulating LH.30 These findings suggest that IGF-I can modulate GnRH neurons by inducing gene expression, resulting in more circulating LH.
Other substances found to affect GnRH and LH secretion include GABA and opioids. GABA may tonically inhibit LH secretion, as evidenced by an increase in LH secondary to a GABA-A receptor blocking drug, bicuculline.52 Naloxone is an opioid antagonist that has been shown to increase luteal phase LH, suggesting that opiates have an inhibitory effect on pituitary gonadotropins.53
Hyperandrogenemia
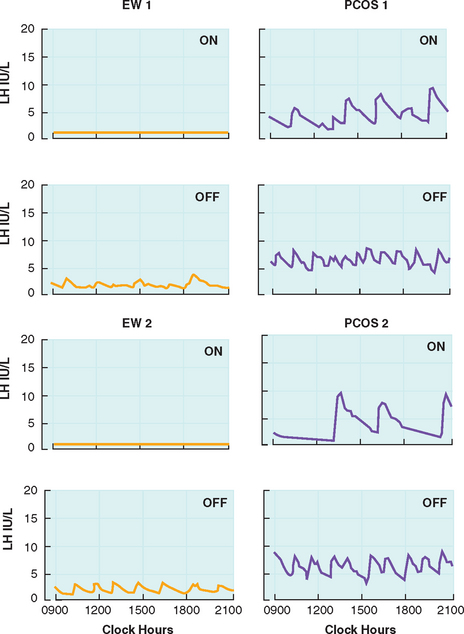
Circulating androgens are elevated in PCOS, with contributions from the ovary and adrenal glands. The elevated androgens can only be partially suppressed with combination oral contraceptive therapy. Daniels and Berga treated PCOS women with 3 weeks of combination oral contraceptives and found that androstenedione levels remained significantly higher compared to treated controls.40 Pulse frequency of LH was suppressed in both PCOS women and controls, but the frequency remained significantly higher in PCOS patients (Fig. 15-3). This suggests reduced sensitivity of the GnRH pulse generator to suppression by sex steroids. The authors also suggest that the GnRH drive in PCOS women may be intrinsically and irreversibly faster than in eumenorrheic women. GnRH output may be affected by peripheral androgens. Androgen blockade restores hypothalamic sensitivity to feedback inhibition by ovarian steroids.54 Administration of estrogen and progesterone to PCOS women and controls pretreated with the anti-androgen flutamide demonstrated similar reductions in LH pulse frequency.
Theca Cell Function
Ovarian hyperandrogenism is driven by LH acting on theca cells, and the effect is amplified by the increased sensitivity of PCOS theca cells to LH.55 Hyperandrogenism may also result from dysregulation of the androgen-producing enzyme P450c17, which has 17α-hydroxylase and 17,20-lyase activity. The dysregulation appears to be an intrinsic abnormality of P450c17, but autocrine/paracrine factors may also be involved.56
Insulin and IGF have also been shown to play a role in ovarian androgen production. Receptors for insulin, IGF-I, and IGF-II have been localized to the theca compartment of ovaries from normal and PCOS patients.57,58 When given IGF-I, IGF-II, and insulin, thecal cell cultures from euandrogenic women produced increased androgens, and the effect was potentiated when LH was added.57,59 Ovarian stroma from hyperandrogenic patients also released more androstenedione and testosterone in response to insulin, however, no synergy was found between insulin and LH.60
In contrast, in vivo studies do not find significant increases in androgen secretion in PCOS or normal women despite considerable increases in insulin levels. Dunaif and Graf studied PCOS and normal patients undergoing evaluation with the hyperinsulinemic euglycemic clamp, and found that insulin decreased androgens in PCOS women and did not increase androgens in normal women.13 This argues against a simple causal relationship between hyperinsulinemia and hyperandrogenism, but a role for insulin is strongly suggested by the observation that reduction of hyperinsulinemia is associated with decreases in serum androgens. Treatment of PCOS patients with metformin, which reduces hepatic glucose production and secondarily lowers insulin, has been shown to decrease levels of testosterone, dehydroepiandrosterone sulfate (DHEAS), and androstenedione.61
Adrenal Function
Excess adrenal androgen production is seen in PCOS women, with a 48% to 64% increase in DHEAS and 11β-hydroxyandrostenedione.9 The underlying cause of elevated adrenal androgens is yet to be elucidated, but PCOS women do not have increased corticotropin levels.62 Increased adrenal androgen production in PCOS is likely caused by either altered adrenal responsiveness to corticotropin or abnormal adrenal stimulation by factors other than corticotropin. A recent study by Moran and colleagues found that adrenal androgen excess in PCOS is associated with a greater 17α-hydroxylase activity in response to corticotropin.63 The cytochrome P450c17 (CYP17) gene regulates 17α-hydroxylase and 17,20-lyase activity. A defect in the P450c17 enzyme or in the regulation by autocrine/paracrine factors appears to be involved.56
Hyperinsulinemia may also play a role in adrenal androgen production in PCOS. PCOS patients treated with troglitazone experienced improvement in insulin resistance with a concomitant decrease in DHEAS levels, regardless of initial DHEAS levels.64 Obese PCOS women treated with pioglitazone had significant improvement in insulin sensitivity with a decrease in corticotropin-stimulated androstenedione levels.65 These data support the notion that insulin enhances corticotropin-stimulated steroidogenesis.
Anovulation
Granulosa Cell Function
FSH levels are characteristically low in PCOS women, resulting in arrested follicular development. Insufficient granulosa cell aromatase activity was the basis of earlier studies that tried to explain poor follicular development, because follicular fluid estradiol concentrations were thought to be low. To the contrary, more recent studies found that PCOS granulosa cells are hyperresponsive to FSH in vitro, and estradiol concentrations from PCOS follicles and normal follicles are no different.66 A dose–response study in PCOS women demonstrated a significantly greater capacity for estradiol production in response to recombinant human FSH compared with normal women.67 The incremental response of serum estradiol was almost two times greater and considerably accelerated compared with that found in normal women.
Van Der Meer and coworkers demonstrated that the accelerated estradiol production may simply be explained by the increased number of follicles.68 Another study of granulosa cells from polycystic ovaries discovered that there are more FSH receptors compared with cells from normal ovaries.69 Whether the increased estradiol responsiveness to FSH is due to increased number of follicles, increased granulosa cell sensitivity to FSH, or increased FSH receptor binding, women with PCOS appear to be susceptible to ovarian hyperstimulation in response to gonadotropins.
Insulin has been investigated as a modulator of both theca and granulosa cell function. As noted earlier, insulin has been shown to augment the effect of LH on thecal cell androgen secretion. The effects of insulin on granulosa cells are less well known. In vitro studies have shown that insulin augments two actions of FSH on granulosa cells: basal production of estrogen and progesterone and induction of LH responsiveness.70,71 The latter effect results in further enhanced LH-stimulated steroid production of estradiol and progesterone. Willis and Franks showed that the effects of insulin were apparent at low concentrations, underscoring the fact that insulin acts via its own receptor and suggesting that the ovary is not insulin resistant.72 The premature responsiveness of granulosa cells to LH may lead to untimely differentiation of granulosa cells, resulting in premature arrest of follicular growth. This might explain the presence of the many 5- to 10-mm follicles commonly observed in PCOS ovaries containing steroidogenically active granulosa cells that do not progress spontaneously to the preovulatory stage.
Insulin Resistance
Although 50% to 70% of PCOS patients have insulin resistance,73 it is not one of the diagnostic criteria for PCOS. The topic deservedly receives much attention because many of the clinical signs and symptoms of PCOS may be attributed to excess insulin exposure. The precise molecular basis for insulin resistance is unknown, but it appears to be a postreceptor defect.74 There is tissue specificity of insulin resistance in PCOS: muscle and adipose tissue are resistant; the ovaries, adrenals, liver, skin, and hair remain sensitive. The resistance to insulin in skeletal muscle and adipose tissue leads to a metabolic compromise of insulin function and glucose homeostasis, but there is preservation of the mitogenic and steroidogenic function in other tissues. The effect of hyperinsulinemia on the sensitive organs results in the downstream effects seen in PCOS, such as hirsutism,7 acanthosis nigricans,17 obesity,14 stimulation of androgen synthesis,57 increase in bioavailable androgens via decreased sex hormone-binding globulin (SHBG),75 and, potentially, modulation of LH secretion.60
In 1992, Hales and Barker proposed the concept that the environmental influence of undernutrition in early life increased the risk of type 2 diabetes in adulthood.76 They discovered a relationship between low birth weight and type 2 diabetes in men from England. The finding has been replicated in many different populations and ethnic groups, and the relationship has been extended to include the antecedent condition of insulin resistance.77 In the thrifty phenotype hypothesis, malnutrition serves as a fetal and infant insult that results in a state of nutritional thrift. The adaptations result in postnatal metabolic changes that prepare the individual for survival under poor nutritional conditions. One advantageous change is the development of insulin resistance in muscle and adipose tissue, with selective protection of brain growth and activity. The adaptations become detrimental when the postnatal environment changes to one of an overabundance of nutrients, resulting in obesity and diabetes.
Recognition of insulin resistance can identify those who are most likely to respond to lifestyle and pharmacologic intervention. Insulin resistance puts patients at increased risk for certain metabolic sequelae, such as diabetes, hypertension, dyslipidemia, and cardiovascular disease.78 Insulin resistance is a component of the World Health Organization (WHO) definition of the metabolic syndrome, which is a cluster of risk factors for cardiovascular disease.79 The WHO defines the metabolic syndrome as the presence of glucose intolerance or insulin resistance, with at least two of the following: hypertension, dyslipidemia, obesity, and microalbuminuria. Women with PCOS are 4.4 times more likely to have the metabolic syndrome, so it becomes prudent to screen these patients, especially in those with insulin resistance.80






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


