Carboxylic acids
Pyrazoles
Oxicams
COX-2 selective
Salicylates
Acetic acids
Propionic acids
Fenamates
Acetylsalicylic acid salsalate diflunisal fendosal
Indomethacin acemetacin cinmetacin sulindac tolmetin zomepirac diclofenac fenclofenac isoxepac furofenac fentiazac clidanac oxepinac fenclorac lonazolac metiazinic acid clopirac amfenac benzofenac clometacine etodolac bumadizone clamidoxic acid
Ibuprofen flurbiprofen naproxen ketoprofen fenoprofen benoxaprofen indoprofen pirprofen carprofen oxaprozin pranoprofen suprofen miroprofen tioxaprofen alminoprofen cicloprofen tiaprofenic acid furaprofen butibufen fenbufen furobufen bucloxic acid protizinic acid
Mefenamic acid flufenamic acid meclofenamate niflumic acid toifenamic acid flunixin clonixin
Phenyibutazone feprazone apazone trimethazone mofebutazone kebuzone suxibuzone
Piroxicam isoxicam tenoxicam
Celecoxib rofecoxib valdecoxibe nimesulide NS 398
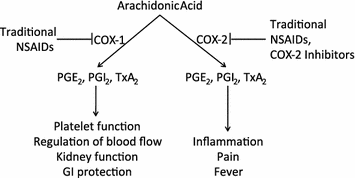
Fig. 1
Prostaglandins and thromboxanes produced through COX-1 have important physiological functions, whereas the prostaglandins and thromboxanes produced through COX-2 have important pathophysiological functions
The specific biochemical and cellular mechanism(s) proposed to be responsible for the cancer chemopreventive activity of the NSAIDs is controversial. While there is strong evidence that the mechanism of action involves COX-2 inhibition, there are a number of pharmacological inconsistencies that have led many investigators to conclude that the mechanism is unrelated to COX-2 inhibition (Alberts et al. 1995; Elder et al. 1997; Hanif et al. 1996; Kashfi and Rigas 2005; Piazza et al. 1997a). For example, studies have shown that NSAIDs can inhibit the growth of tumor cells that completely lack the expression COX-2 (Elder et al. 1997; Hanif et al. 1996; Grosch et al. 2001). Other studies have reported that the addition of exogenous prostaglandins cannot rescue cancer cells from the growth inhibitory activity of the NSAIDs, which would be expected if prostaglandin suppression is necessary (Kusuhara et al. 1998; Piazza et al. 1995). Furthermore, as shown in Table 2, the rank order of potency among the NSAIDs to inhibit tumor cell growth does not correlate with their potency to inhibit prostaglandin synthesis (Carter et al. 1989; de Mello et al. 1980; Erickson et al. 1999). For example, sulindac sulfide is appreciably less potent than indomethacin with regard to COX-1 or COX-2 inhibition, but is more potent in terms of its tumor cell growth inhibitory activity (Tinsley et al. 2010). In addition, certain highly specific COX-2 inhibitors such as rofecoxib fail to inhibit tumor cell growth (Soh et al. 2008). Consequently, a number of alternative targets have been suggested as potential mediators (Chan 2002; Thun et al. 2002; Shiff and Rigas 1999).
Table 2
Potencies of select NSAIDs and COX-2 selective inhibitors for inhibition of HT-29 colon tumor cell growth, inhibition of purified ovine COX-2, and inhibition of cGMP PDE activity in HT-29 cell lysate
IC50 (μM) | |||
|---|---|---|---|
Growth inhibition | COX-2 inhibition | cGMP PDE inhibition | |
Tolmetin | 313 | 0.82 | 326 |
Meclofenamic acid | 85 | 2.9 | 80 |
Flufenamic acid | 108 | 9.3 | 103 |
Flurbiprofen | 550 | 5.5 | 504 |
Celecoxib | 203 | 0.34 | 174 |
Sulindac | 224 | >300 | 120 |
Sulindac sulfide | 34 | 6.3 | 20 |
Sulindac sulfone | 89 | >300 | 96 |
In this review, we consider several important developments in the study of both COX-2-dependent and COX-independent targets that can guide efforts in drug discovery and development strategies for cancer chemoprevention. We also discuss several promising novel NSAID derivatives that have been identified in recent years, which have potential safety and efficacy advantages compared to currently available NSAIDs and COX-2 inhibitors.
2 Targeting COX-2
There is strong evidence for the involvement of COX-2 in colon tumorigenesis. For example, COX-2 is overexpressed and prostaglandin levels are elevated in as many as 90 % of sporadic colon carcinomas (Eberhart et al. 1994). Furthermore, the levels of COX-2 expression correlates with colon tumor size and invasiveness (Fujita et al. 1998). The mechanism by which COX-2 may drive tumorigenesis is still unclear but likely involves multiple pathways given that prostaglandins can accelerate cellular growth, inhibit apoptosis, and induce angiogenesis; all key events in carcinogenesis (Shiff and Rigas 1999; Sawaoka et al. 1998; Tsujii et al. 1998). No matter the mechanism, studies have shown that decreasing COX-2 expression in the Apcmin mouse model of intestinal carcinogenesis reduces both the size and multiplicity of intestinal polyps (Oshima et al. 1996). Similarly, clinical studies have shown efficacy of celecoxib and rofecoxib, albeit modest, for suppressing the formation of colorectal adenomas in patients with familial adenomatous polyposis as well as the formation of sporadic adenomas (Steinbach et al. 2000; Arber et al. 2006; Baron et al. 2006; Bertagnolli et al. 2006).
While the COX-2 selective inhibitors, rofecoxib and valdecoxib, were removed from the market due to unforeseen cardiovascular toxicity, it is unclear if these side effects are directly linked with COX-2 inhibition, especially since they were less pronounced with celecoxib (Chakraborti et al. 2010). Given that the toxicity may be unrelated to COX-2 but may be compound specific, it is possible that chemical modifications to existing NSAIDs might have potential advantages. Marnett and colleagues, for example, described a series of ester and amide modifications to the carboxylic acid moiety on various NSAIDs, such as indomethacin and meclofenamic acid that increased selectivity for COX-2 (Kalgutkar et al. 2000; 2002). The rational for this approach was based on molecular modeling studies whereby a more restrictive binding domain was noted within the catalytic region of the COX-1 compared with COX-2, which permitted specificity via substitution of the carboxylic acid with bulky neutral residues. A potential concern, however, is that ester and amide linkages may not have sufficient metabolic stability whereby there is the potential to generate the parent NSAID. Studies to demonstrate in vivo antitumor efficacy and reduced toxicity compared with the parent NSAID are therefore needed for future development of this class of compounds.
3 COX-Independent Targets
3.1 Inhibition of cGMP PDEs
One COX-independent target of NSAIDs that has been used to guide the synthesis of more potent non-COX inhibitory derivatives includes members of the cyclic GMP phosphodiesterase (PDE) superfamily. PDE enzymes are metallophosphohydrolases that hydrolyze the 3′,5′-cyclic phosphate on second messenger cyclic nucleotides, cAMP or cGMP, to 5′-monophosphate. This modification terminates intracellular signaling following activation of cyclic nucleotide coupled receptors (Beavo 1995). The PDE superfamily comprises an estimated 100 distinct protein isoforms divided into 11 protein families, which differ from one another in selectivity for cAMP or cGMP, sensitivity to inhibitors and activators, as well as tissue, cellular, or intracellular distributions (Beavo 1995; Sonnenburg and Beavo 1994). Inhibition of PDE would result in an increase in the magnitude and/or duration of the cAMP and/or cGMP signal depending on the isozyme selectivity of the inhibitor. Increased intracellular cyclic nucleotide levels can activate specific signaling pathways, which, in the case of cGMP, can lead to activation of cGMP-dependent protein kinase (PKG), cyclic nucleotide-gated ion channels, or certain cGMP binding PDEs; all of which play important roles in regulating cellular activity (Beavo 1995; Lincoln and Cornwell 1993).
Studies have suggested that the cGMP pathway may be perturbed during colorectal tumorigenesis. For example, human colon tumors have been shown to express high levels of the cGMP-specific PDE5 isozyme (Tinsley et al. 2010). In addition, Shailubhai and colleagues demonstrated that mRNA levels for the membrane-associated guanylyl cyclase agonist, uroguanylin, are reduced in both colon adenomas and adenocarcinomas. Interestingly, oral administration of uroguanylin inhibited tumor formation in the Apcmin mouse model of intestinal tumorigenesis, which was associated with increased apoptosis rates within the tumors (Shailubhai et al. 2000). These findings are consistent with the observations that certain cGMP phosphodiesterases may be elevated in colon tumors, leading to reduced cGMP levels, which is necessary to limit tumor cell survival or suppress proliferation. Despite the evidence for a role of cGMP in colon cancer development, the relevant cGMP PDE isozymes that regulate its synthesis and downstream signaling pathways have not been well studied with regard to a role in colon tumorigenesis.
Certain NSAIDs and COX-2 inhibitors can also inhibit cGMP PDE isozymes to increase intracellular cGMP levels and activate cGMP signaling in colon cancer cell lines, as depicted in Fig. 2 (Soh et al. 2000, 2008; Silvola et al. 1982; Thompson et al. 2000; Tinsley et al. 2009). While these effects require high concentrations (Table 2) that cannot be safely achieved in vivo, a strong correlation exists between concentrations of the NSAIDS required to inhibit tumor cell growth in vitro and their ability to inhibit cGMP PDE, especially the cGMP-specific PDE isozyme, PDE5 (Tinsley et al. 2010). In addition, certain known PDE5-specific inhibitors (e.g. MY5445) and non-selective cGMP PDE inhibitors (e.g. MY5445, trequinsin) could also suppress tumor cell growth, as well as knockdown of PDE5 by siRNA or antisense (Tinsley et al. 2011; Zhu et al. 2005). However, other highly selective PDE5 inhibitors (e.g. sildenafil), commonly used for the treatment of erectile dysfunction, required high concentrations compared with those required to inhibit tumor cell growth, which suggest that additional cGMP PDE isozymes may be involved (Tinsley et al. 2011).
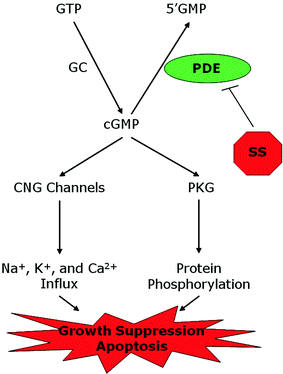
Fig. 2
Mechanistic model of colon tumor cell growth inhibition mediated by inhibition of cGMP PDE and activation of cGMP signaling
Among a number of NSAIDs evaluated for cGMP PDE inhibition, sulindac sulfide was found to be the most active (Tinsley et al. 2010). Since the non-COX inhibitory sulfone form of sulindac can also inhibit cGMP PDE, albeit with less potency, it is likely that the COX inhibitory activity of this class of compounds can be uncoupled from cGMP PDE inhibitory activity (Piazza et al. 1995, 1997b). Initial efforts to synthesize sulindac derivatives that lack COX inhibitory activity, but have improved cGMP PDE inhibitory activity focused on modifications to either the sulfonyl or the carboxylic acid moieties (Thompson et al. 2000). While this approach yielded certain derivatives with high in vitro potency to inhibit colon tumor cell growth and induce apoptosis, poor oral bioavailability and metabolic instability halted further development (Thompson et al. 2000).
In light of information gained from molecular modeling docking sulindac sulfide into the COX active site, our laboratory has focused on chemically modifying the carboxylic acid moiety of sulindac sulfide and substituting with a positive-charged amide to interfere with COX binding (Piazza et al. 2009). This approach yielded an interesting series of compounds that retained ability to inhibit cGMP PDE and had improved anticancer activity both in vitro and in vivo compared with sulindac (Piazza et al. 2009). These derivatives serve to validate cGMP PDE as a target to optimize for anticancer efficacy, while reducing toxicity. Although problems with poor absorption following oral administration necessitated high dosages limited the development of this class of compounds for colorectal cancer chemoprevention, alternative derivatives or formulations with improved oral bioavailability may have advantages.
3.2 Generation of Reactive Oxygen Species
Reactive oxygen species (ROS) such as the superoxide anion or nitric oxide have long been recognized as by-products of mitochondrial oxygen metabolism that can be toxic to cells (Genestra 2007). Furthermore, ROS generated by certain organelles such as peroxisomes, especially in cellular types such as macrophages, mediate important physiological processes including macromolecular anabolism and pathogen destruction (Genestra 2007; Klaunig and Kamendulis 2004). However, recent evidence suggests that these highly reactive molecules can serve as signaling molecules within cells (redox signaling), interacting with numerous canonical signaling pathways to mediate a variety of cellular functions including proliferation, apoptosis, and differentiation (Frein et al. 2005).
Aberrant redox signaling has been documented in cancer, although more work is necessary to determine whether ROS are a driving force in tumorigenesis. For example, abnormally high levels of ROS have been observed in multiple types of cancer (Toyokuni et al. 1995). Furthermore, chronic inflammation, which can promote tumorigenesis, results in elevated levels of ROS. High ROS levels have been reported in tumors and are associated with genomic instability, proliferation, angiogenesis, and aberrant apoptosis, although the effect may be dependent on intracellular levels (Frein et al. 2005; Toyokuni et al. 1995; Halliwell 2007).
Two promising new classes of NSAID derivatives, the nitric oxide NSAIDs (NO-NSAIDs) and the phospho-NSAIDs, demonstrate chemopreventive efficacy through a mechanism that appears to involve the elevation of intracellular ROS levels and activation of proapoptotic redox signaling pathways as described by Rigas and colleagues. The NO-NSAIDs were designed with an NO-releasing moiety attached to a parent compound such as aspirin, sulindac, or naproxen through a chemical linker. The intended design was to increase NO levels in the gut where it could serve a protective function to increase vascular blood supply to reduce the GI toxicity of the parent NSAID (Rigas and Kashfi 2004). Initial studies with the NO-NSAIDs showed enhanced colon cancer chemopreventive activity when compared to the parent compound both in vitro and in vivo (Williams et al. 2004; Yeh et al. 2004). In order to identify the importance of the NO moiety for the enhanced chemopreventive activity, additional modifications were made in which the NO was replaced with a diethylphosphate. Interestingly, these phospho-NSAID derivatives demonstrated similar chemopreventive activity as the NO-NSAIDs, suggesting that release of the NO was not solely responsible for the enhanced activity of these new classes of compounds, but rather a chemical modification to the NSAIDs (Huang et al. 2010; Mackenzie et al. 2010).
Whether the parent NSAID was linked to NO or to the diethylphosphate substituent, the new derivatives caused substantial increases in ROS in tumor cells in vitro and showed advantages in efficacy in animal models of colon cancer (Rigas and Kashfi 2004; Kashfi and Rigas 2007; Rigas and Kozoni 2008; Rigas and Williams 2008; Sun et al. 2011; Xie et al. 2011). While further studies are necessary to develop this class of derivatives, their ability to induce apoptosis of colon tumor cells through activation of redox signaling makes them a promising class of agents for colorectal cancer chemoprevention.
3.3 Downregulation of Survivin
Survivin is an apoptosis inhibitor protein that prevents activation of caspases that are important mediators of apoptosis (Altieri 2003a, b; Ambrosini et al. 1997). Not normally found to be expressed in adult tissues, survivin is overexpressed in the vast majority of human cancers and increased levels of this protein strongly correlate with increased tumor stage and a poor prognosis (Altieri 2003a; Ambrosini et al. 1997; Zaffaroni et al. 2005). Furthermore, survivin expression appears to be strongly connected to p53 status and sensitivity to chemotherapy (Hoffman et al. 2002; Li 2005).
A number of NSAIDs and COX-2 selective inhibitors have been shown to reduce survivin expression and/or activity in cancer cells, but the best studied of these is celecoxib (Tinsley et al. 2010, 2011; Konduri et al. 2009; Pyrko et al. 2006). Interestingly, the celecoxib derivative 2,5-dimethyl-celecoxib (DMC) lacks COX-2 inhibitory activity but retains the anticancer activity of the parent compound and is actually more potent than celecoxib for suppressing survivin (Pyrko et al. 2006). Even more promising, DMC has shown in vivo chemopreventive efficacy in various models of human cancer (Pyrko et al. 2006; Kardosh et al. 2005).
3.4 Other COX-Independent Targets
Additional targets that are known to play a role in colorectal cancer tumorigenesis have also been implicated as targets of NSAIDs. However, the association between these targets and the anticancer activity of the NSAIDs is weaker and has not been established for non-COX-inhibitory derivatives, but nonetheless represent potential targets for future efforts in drug discovery.
One such target is peroxisomal proliferator-activated receptor γ (PPARγ). PPARγ is overexpressed in colon cancer cells and can inhibit growth and promote differentiation (DuBois et al. 1998; Mueller et al. 1998). Numerous studies have shown that certain NSAIDs can directly bind to and activate PPARγ and that this activity is associated with inhibition of neoplastic growth (Lehmann et al. 1997). However, the activation of PPARγ is not a characteristic shared by all antineoplastic NSAIDs (Lehmann et al. 1997; Lefebvre et al. 1998; Saez et al. 1998).
NFκB is another potential COX-independent antineoplastic target of the NSAIDs. NFκB is a transcription factor that commonly promotes cellular growth and inhibits apoptosis (Ljungdahl et al. 1997). Increased NFκB activity has been observed in multiple types of cancer and can be attributed to a number of alterations including overexpression of NFκB or decreased expression of IκB, a regulatory protein that sequesters NFκB in the cytosol in order to prevent its transcriptional activity (Rayet and Gelinas 1999). Some NSAIDs, particularly the salicylates, have been found to inhibit NFκB-mediated transcription, presumably by preventing degradation of IκB (Schwenger et al. 1998; Yin et al. 1998). However, these effects are most pronounced among the least active NSAIDs in terms of in vitro colon cancer prevention (Yin et al. 1998).
One of the most novel mechanisms proposed for the anticancer activity of the NSAIDs involves induction of the NSAID-activated gene (NAG-1) as reported by Baek and colleagues. NAG-1 is a member of the transforming growth factor β (TGF-β) superfamily that has both proapoptotic and antiproliferative properties, although the exact mechanism responsible for these effects has not been well defined (Baek et al. 2001a, b). A number of different NSAIDs including indomethacin, aspirin, and ibuprofen have been shown to increase expression of NAG-1 in colon cancer cells and this effect appears to be independent of COX expression (Baek et al. 2001a, 2002). However, there is little association between the anticancer activity of an NSAID and its potency for activating NAG-1 expression, as NAG-1 induction occurs at significantly lower concentrations than those necessary for induction of apoptosis or inhibition of growth (Baek et al. 2002). Furthermore, the effects of the NSAIDs on NAG-1 and the effects of NAG-1 expression have yet to be observed in vivo.
Although not a direct target, microRNA may be another important factor that contributes to the sensitivity of tumor cells to NSAIDs. microRNAs are a set of naturally occurring small RNA molecules that are capable of regulating approximately 30 % of human genes through direct binding with the cognate target genes and are involved in many essential biological processes such as cell growth, differentiation, apoptosis, and tumorigenesis (Carmell et al. 2002; Esquela-Kerscher and Slack 2006), which highlights its clinical applications in tumor diagnosis, prognosis, and therapy (Nakajima et al. 2006; Xi et al. 2006). Our studies, for example, have shown that a panel of microRNAs (miR-10b, -17, -19, -21, and -9) are suppressed in human colon tumor cells treated with sulindac sulfide (Rayet and Gelinas 1999). Interestingly, these microRNAs are known to be elevated during cancer metastasis and invasion (Huang et al. 2009; Ma et al. 2010a, b, 2007; Song et al. 2010; Yu et al. 2010; Zhu et al. 2008) and may mediate the ability of sulindac sulfide to inhibit tumor cell invasion.
Another complicating aspect of determining the mechanisms of the actions of NSAIDs in the prevention of neoplasia is the effects of various types of NSAIDs on inflammation in general. Inflammation in the setting of long standing, continuing damage, inflammation, and repair that occurs in ulcerative colitis and the associated development of colon cancers generates not only increased ROS, but also increased cellular death and proliferation. These associated inflammatory changes may lead to molecular changes in epithelial cells that result in the inhibition of enzymes that repair DNA to molecular changes in signaling pathways associated with the initiation of neoplasia as a result of genetic mutations (Baek et al. 2001b, 2002). Thus, some changes related to the chemopreventive actions of NSAIDs may be related specifically to their anti-inflammatory activity involving COX-inhibition, while others (e.g. tumor cell growth inhibition and apoptosis induction) are more related to their ability to suppress tumorigenesis. Whether the two activities can be fully uncoupled remains to be determined and awaits the development of new derivatives that lack COX inhibitory activity.
4 Conclusions
NSAIDs represent a chemically diverse group of drugs that have multiple biological effects, some of which are related to their anticancer activities, while others are responsible for toxicity. Experimental studies over the past 20 years have provided strong evidence that the mechanism responsible for their chemopreventive activity may not require COX inhibition. As such, it should be feasible to develop improved drugs with greater antitumor efficacy and reduced toxicity. Given that the anticancer activity of the NSAIDs is undoubtedly complex, a number of reports have suggested many alternative targets as summarized in Table 3. Traditional NSAIDs and COX-2 selective inhibitors most likely inhibit tumorigenesis through a combination of COX-dependent and COX-independent mechanisms. However, the development of NSAID derivatives that lack COX inhibitory activity have shown promise for improved potency and selectivity to inhibit tumor cell growth. As such, the elucidation of COX-independent mechanisms of the NSAIDs is an important area of research that offers the promise to design a highly specific new class of chemopreventive agents. The greatest challenges will be to identify which molecular target(s) is most important for colon tumorigenesis and which chemical scaffold can yield suitable lead compounds for preclinical development with optimal pharmaceutical, efficacy, and toxicity properties.
Table 3
 Emerging Role for Anti-inflammatory Agents for Chemoprevention
Agents with Anti-lnflammatory Properties in Chemoprevention of Colorectal Neoplasia
Emerging Role for Anti-inflammatory Agents for Chemoprevention
Agents with Anti-lnflammatory Properties in Chemoprevention of Colorectal Neoplasia
 Aspects of COX-2 Expression in Colorectal Neoplasia
Aspects of COX-2 Expression in Colorectal Neoplasia
 Pharmacology, Toxicity and Efficacy in Cancer Clinical Trials
Pharmacology, Toxicity and Efficacy in Cancer Clinical Trials
 Inheritance and Strategies for Prevention in Populations at High Risk of Colorectal Cancer (CRC)
Inheritance and Strategies for Prevention in Populations at High Risk of Colorectal Cancer (CRC)
 in Prevention of Sporadic Colorectal Cancer: Current Clinical Evidence and Overall Balance of Risks and Benefits
in Prevention of Sporadic Colorectal Cancer: Current Clinical Evidence and Overall Balance of Risks and Benefits




Summary of drugs and COX-independent targets that have been studied for the chemopreventive activity of the NSAIDs and COX-2 selective inhibitors
Related posts:
Emerging Role for Anti-inflammatory Agents for Chemoprevention
Agents with Anti-lnflammatory Properties in Chemoprevention of Colorectal Neoplasia
Aspects of COX-2 Expression in Colorectal Neoplasia
Pharmacology, Toxicity and Efficacy in Cancer Clinical Trials
Inheritance and Strategies for Prevention in Populations at High Risk of Colorectal Cancer (CRC)
in Prevention of Sporadic Colorectal Cancer: Current Clinical Evidence and Overall Balance of Risks and Benefits
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
