Chapter 4 Liver blood flow
Physiology, measurement, and clinical relevance
Physiology
Liver Blood Supply
The liver normally receives about one quarter of the total cardiac output despite constituting only 2.5% of total body weight; it obtains its blood supply from two main sources, the hepatic artery and the portal vein. Approximately 75% to 80% of the blood entering the liver is partially deoxygenated portal venous blood drained from the stomach, intestine, spleen, and pancreas. The remainder is well-oxygenated blood from the aorta, carried by the hepatic artery. Mixing of arterial and portal blood occurs in the sinusoids, which are drained by the hepatic venous system into the inferior vena cava. Hepatic blood volume accounts for 10% to 15% of total blood volume with 40% of hepatic blood held in large vessels such as the hepatic arteries, portal vein, and hepatic veins; the remaining 60% is held in the sinusoids (Lautt, 1977; Greenway & Stark, 1971).
Hepatic Artery
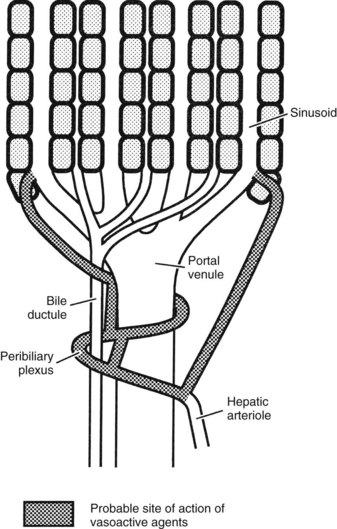
The hepatic arterioles empty directly or via a peribiliary plexus into the sinusoids and terminal portal venules. Direct artery-to-hepatic vein connections do not usually exist but may arise in liver disease. Reduction of pressure in the arterial system toward that existing in the portal circulation is achieved mainly by 1) the presinusoidal arteriolar resistance in the peribiliary plexus and 2) the intermittent closure of the arterioles, which protect the portal bloodstream from arterial pressure (Rappaport, 1973).
In the event of hepatic arterial occlusion, numerous small arteries provide a source for the formation of a collateral circulation. Upon interruption of hepatic arterial flow, collaterals can arise from 1) the inferior phrenic arteries, which can develop connections with hepatic arteries within the liver, and 2) from the gastroduodenal arteries, which derive blood flow from the superior mesenteric artery and supply the liver via the peribiliary arterial plexus around the intrahepatic bile ducts (Rappaport & Schneiderman, 1976). The precise nature of the functional collateral supply after hepatic arterial ligation depends on the site of occlusion. If the common hepatic artery is interrupted, revascularization occurs through both major routes; if only the right or left hepatic artery is interrupted, intrahepatic translobar anastomoses reestablish arterial flow in the ligated system (Mays & Wheeler, 1974). Ligation of the proper hepatic arteries leads to revascularization mainly via a hypertrophied inferior phrenic circulation (Jefferson et al, 1956). Complete long-term dearterialization of the liver by any form of arterial vascular occlusion is difficult to achieve.
Portal Vein
The inferior and superior mesenteric veins join with the splenic vein to form the portal vein, and jointly they collect the venous outflow from the entire prehepatic splanchnic vascular bed (the intestinal tract from the lower esophagus to the rectum plus the pancreas and spleen). This valveless system is a low-pressure/low-resistance system regulated by the degree of constriction of mesenteric and splanchnic arterioles coupled with the intrahepatic vascular resistance. The portal vein normally carries about 75% of the total blood flow to the liver, or 90 mL/min per 100 g of liver weight; normal portal pressure is 5 to 8 mm Hg. Portal blood is postcapillary and partly deoxygenated, but because of its large volume flow rate, it may supply 50% to 70% of the liver’s normal oxygen requirement. During fasting states, the oxygen saturation in the portal blood approaches 85%, which is greater than other systemic veins. Hepatic oxygen supply is diminished if portal blood flow is significantly reduced, but the effect is minimized by an increase in oxygen extraction from the hepatic arterial blood and not by increasing flow. In fact, isovolemic hemodilution or upregulation of hepatic enzymes leading to oxygen deprivation does not result in hepatic artery dilatation, refuting the concept that arterial flow is regulated by liver cell metabolism (Lautt et al, 1987).
Hepatic Veins
The hepatic venous system is the systemic drainage tract of the entire splanchnic circulation. A total liver blood flow (LBF) of 1.5 L/min is considered the normal value in the average male, but the range can be quite wide (1 to 2 L/min). Although the free pressure in a hepatic vein is 1 to 2 mm Hg, the wedged hepatic venous pressure, which is useful for estimating sinusoidal pressure, may be an indicator of portal venous pressure (Boyer et al, 1977). The sinusoidal pressure is estimated to be slightly higher than that of the vena cava, or 2 to 4 mm Hg (Eipel et al, 2010). Hepatic venous blood is normally about two-thirds saturated with oxygen, but this may be markedly reduced during periods of low delivery of oxygen to the liver, when oxygen is extracted by hepatocytes. In resting states, the liver accounts for approximately 20% of the total oxygen consumption of the body.
Control of Liver Blood Flow
Some controversy surrounds the autoregulation of hepatic arterial vasculature. Although some studies have suggested nonlinear pressure-regulated changes in arterial resistance in denervated dogs (Hanson et al, 1966), others have demonstrated a more direct relationship (Torrance, 1961). The pressure-flow dynamic in the portal system has been found to have little to no autoregulation, as evidenced by partial collapse of the portal vascular bed in response to reduced portal pressure (Condon et al, 1962).
Intrahepatic Vascular Resistance in Health
Intrahepatic vascular resistance (IHVR) seems to be located at the level of the portal venules (presinusoidal) or sinusoids (Sherman et al, 1996; Shibayama & Nakata, 1985; Zhang et al, 1994, 1995). Knisley suggested the presence of sphincter-like structures at the entrance and exit of sinusoids that maintain tremendous portal venous pressure gradients, but these may be species dependent (Knisley et al, 1957). In dogs and cats, IHVR is located at the level of hepatic venules (postsinusoidal) under resting conditions (Lautt et al, 1986, 1987), whereas it may be shifted to presinusoidal sites during neural stimulation (Greenway & Lautt, 1970; Legare & Lautt, 1987). Sinusoidal contraction in response to the vasoconstrictor endothelin (ET) 1 has been observed despite the lack of smooth muscle (Bauer et al, 1994, 1995; Okumura et al, 1994; Zhang et al, 1994, 1995). This contractility is mainly attributed to hepatic stellate cells—also called Ito cells, lipocytes, and fat-storing cells—which are distinguished by autofluorescence derived from their intracellular vitamin A (Suematsu et al, 1995; Zhang et al, 1994). ET-1 causes reversible and graded contraction of isolated stellate cells in tissue culture and results in narrowing of sinusoidal lumens in perfused livers (Pinzani et al, 1992; Rockey & Weisiger, 1996; Rockey et al, 1992; Zhang et al, 1994). Hepatic stellate cells have been compared to pericytes, because they contain smooth muscle–specific intermediate desmin-like and actin-like filaments that encircle neighboring sinusoids (Greenwel et al, 1991; Martinez-Hernandez, 1985; Martinez-Hernandez & Amenta, 1993; see Chapter 6).
Studies using intravital fluorescent microscopy showed that the sites of sinusoidal dilation and constriction is colocalized with that of an autofluorescent vitamin A substance (Suematsu et al, 1995, 1996; Zhang et al, 1994, 1995). Hepatic stellate cells acting as liver-specific pericytes (Friedman, 1997; Martinez-Hernandez, 1985; Martinez-Hernandez & Amenta, 1993) may play a crucial role in modulating IHVR and blood flow, especially at the sinusoidal level (Bauer et al, 1994; Okumura et al, 1994; Pannen et al, 1996; Zhang et al, 1994, 1995). Endothelium-derived nitric oxide (NO) in the hepatic sinusoids may modulate portal resistance under physiologic circumstances (Bauer et al, 1997; Shah et al, 1997). Electron microscopy studies have also demonstrated the presence of smooth muscle cells in the sublobar veins (Aharinejad et al, 1997), as well as terminal hepatic venules (Sasse et al, 1976), which may contribute to IHVR.
Relationship between Hepatic Artery and Portal Vein Blood Flow
Individual control of the hepatic arterial and portal venous circulations is complicated by the existence of a complex interaction between the two systems within the liver. Studies have shown an increase in hepatic arterial blood flow following reduction of portal blood flow (Kock et al, 1972; Mathie et al, 1980a; Schenk et al, 1962); however, a compensatory increase in portal flow following decreased arterial perfusion has rarely been observed (Mathie, 1997). Intraoperative measurements of hepatic artery and portal vein flow in anesthetized patients demonstrated a significant increase in hepatic arterial flow, up to 30% of baseline, with temporary occlusion of the portal vein and no effect on portal flow on occlusion of the hepatic artery (Jakab et al, 1995).
The ability of the hepatic artery to respond acutely to changes in portal flow is referred to as the hepatic arterial buffer response (Lautt, 1981). Highlighting its early embryonic origin, in experiments measuring fetal hepatic arterial blood flow, it has been shown that this regulatory process is even present prenatally (Ebbing et al, 2008). Experimental studies support the view that adenosine, accumulating in the liver as a result of reduced hepatic outflow or released from the walls of blood vessels in response to partial tissue hypoxia, has a significant role in the regulation of the buffer response (Lautt & Legare, 1985; Mathie & Alexander, 1990). This adenosine washout hypothesis is supported by the fact that portal infusion of adenosine increases hepatic arterial blood flow by one half to one third the amount of direct arterial instillation (Lautt et al, 1987). The demonstration of receptors mediating dilation to adenosine in the hepatic arterial vascular bed reinforces this view (Mathie et al, 1991; Fig. 4.1).
Metabolism
Hepatic arterial blood flow has not been traditionally linked to liver metabolism (Lautt, 1983; Mathie & Blumgart, 1983a). It has been shown that neither altered oxygen supply nor bile secretion causes a dependent change in arterial flow (Lautt, 1983); instead, the physiologic role of the buffer system is to maintain hepatic clearance in the face of reduced portal flow and to maintain adequate tissue oxygenation. This is despite the fact that the liver receives more oxygen than it requires and can therefore extract more by hepatocyte metabolism if needed. Therefore arterial flow is independent of oxygenation and is secondary to the nutrient and hormonal needs of the liver.
Blood Gas Tensions
Experimental studies have attempted to clarify the role of arterial blood gas tensions and pH on the hepatic circulation. Hypercarbia (Paco2 > 70 mm Hg) increases portal venous flow and decreases hepatic arterial flow in dogs (Hughes et al, 1979a), whereas hypocarbia (Paco2 < 30 mm Hg) decreases both (Hughes et al, 1979b). Systemic hypoxia (Pao2 < 70 mm Hg) causes a decrease in arterial flow but has no effect on the contribution from the portal vein (Hughes et al, 1979c). The response to metabolic acidosis is similar to that induced by hypercarbia, whereas metabolic alkalosis has essentially no significant effect (Hughes et al, 1980b). The sympathetic nervous system is thought to be responsible for the hepatic arterial vasoconstriction observed in hypercarbia and hypoxia (Mathie & Blumgart, 1983b).
Sympathetic Nervous System
Denervation experiments have shown that the sympathetic nervous system is not involved in basal arterial tone in the liver (Mathie & Blumgart, 1983b). Hepatic sympathetic nervous stimulation causes hepatic arterial vasoconstriction and reduced blood flow, but this is secondary to the autoregulatory response (Greenway & Stark, 1971). Sensory denervated rats and pigs have a diminished arterial buffer response on partial occlusion of the portal vein (Ishikawa, 1995). Portal pressure increases as a result of an increase in portal venous resistance, but portal flow does not decrease, unless there is a decrease in intestinal or splenic blood flow caused by simultaneous sympathetic stimulation of these vascular beds. The liver is a significant blood reservoir, and 50% of its blood volume may be mobilized by nerve stimulation (Greenway & Lautt, 1989).
Although the hepatic artery contains both α-adrenergic and β-adrenergic receptors, the portal venous system is believed to contain only α-receptors (Richardson & Withrington, 1981). At low doses, epinephrine causes hepatic and mesenteric arterial vasodilation, whereas at high doses, vasoconstriction occurs in the hepatic arterial and portal venous vascular beds and in the mesenteric circulation (Greenway & Stark, 1971; Richardson & Withrington, 1981).
Other Endogenous Vasoactive Agents
Hepatic blood flow is profoundly increased by glucagon as a consequence of its strong vasodilatory action on the mesenteric vasculature, but insulin has little hemodynamic effect on the hepatic circulation. Histamine causes hepatic arterial dilation and, in the dog only, hepatic venous constriction. Bradykinin is a potent hepatic arterial vasodilator that has little effect on the portal venous system. The hepatic arterial vascular bed is dilated by most prostaglandins; however, prostacyclin does not affect hepatic arterial flow but increases portal blood flow through a vasodilator effect on the prehepatic vascular bed. NO causes vasodilation in the hepatic arterial and mesenteric vascular beds as discussed earlier. Each of the gut hormones—gastrin, secretin, cholecystokinin, and vasoactive intestinal peptide—causes vasodilation of the hepatic artery. In addition, antagonists of calcitonin gene–related peptide and neurokinin significantly reduce hepatic arterial blood flow, suggesting the presence of their receptors on the arterial vasculature (Biernat et al, 2005).
Angiotensin decreases hepatic arterial and portal blood flow and is one of the few substances to produce a significant vasoconstrictor effect on the hepatic artery. However, the ETs that can exert a powerful and prolonged generalized systemic constriction (Miller et al, 1989; Zhang et al, 1994) also have a direct effect on the hepatic blood flow. ETs reduce hepatic perfusion (Kurihara et al, 1992), increase portal pressure (Bauer et al, 1994; Isales et al, 1993; Tanaka et al, 1994; Tran-Thi et al, 1993), and reduce sinusoidal diameter (Bauer et al, 1994; Okumura et al, 1994; Zhang et al, 1994). Vasopressin decreases portal flow and pressure by mesenteric arterial vasoconstriction but has variable effects on the hepatic artery. Serotonin is believed to mediate vasoconstriction of portal radicles and has been targeted as a substance involved in the maintenance of portal hypertension, which will be discussed later. In contrast, hydrogen sulfide either endogenously or exogenously can reverse the norepinephrine-induced vasoconstriction in an NO-independent fashion (Fiorucci et al, 2005). Intraportal administration of exogenous vasoactive agents affects hepatic arterial resistance (Lautt et al, 1984); the mechanisms underlying this intrahepatic transvascular effect are not understood, but it is likely that the close anatomic association between arterioles and venules (see Fig. 4.1) could permit this and may be a means by which hepatic arterial blood flow is finely controlled by endogenous agents such as gut hormones.
Anesthetic Agents
Hepatic arterial and portal venous blood flow decreases passively in parallel with cardiac output during halothane inhalation, with little change in vascular resistance (Hughes et al, 1980a; Thulin et al, 1975). Hepatic oxygen consumption is not diminished by halothane because of a marked increase in the oxygen extraction rate from the reduced blood supply (Andreen et al, 1975). Enflurane has been found to have similar effects to those of halothane, although there is a decrease in hepatic arterial vascular resistance as part of a generalized decrease in peripheral vascular resistance (Hughes et al, 1980a). Cyclopropane and methoxyflurane reduce LBF, mainly by increasing the mesenteric vascular resistance (Batchelder & Cooperman, 1975). NO in concentrations of 30% to 70% reduces hepatic artery and portal vein flow, possibly as a result of a generalized stimulatory action on α-adrenergic receptors (Thomson et al, 1982). Isoflurane seems to have minimal effects on hepatic arterial and portal venous flows, and the intravenous agent fentanyl may have little effect on prehepatic splanchnic blood flow (Nagano et al, 1988); thiopentone in low doses vasoconstricts the hepatic arterial and mesenteric vascular beds (Thomson et al, 1986).
Measurement of Liver Blood Flow
The earliest methods of measuring LBF involved direct invasive techniques, such as intravascular devices or venous outflow collection (Burton-Opitz, 1910, 1911; MacLeod & Pearce, 1914). Indirect determination of blood flow by the use of a variety of indicator-clearance techniques were subsequently developed but were confounded by the presence of liver disease. The available methods are discussed under three broad headings: 1) flow in single blood vessels, 2) total LBF, and 3) hepatic tissue perfusion. The techniques currently employed for experimental and clinical investigations are listed in Table 4.1.
Table 4.1 Summary of Methods Currently Used for Measuring Liver Blood Flow
| Flow in Single Vessels |
| Total Liver Blood Flow |
| Hepatic Tissue Perfusion |
Flow in Single Vessels
Electromagnetic Flowmeter
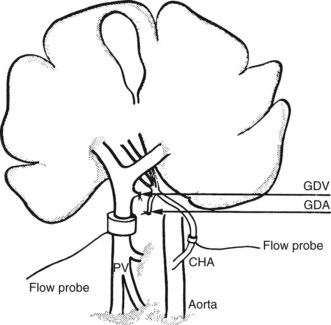
The direct and continuous measurement of hepatic arterial and portal venous blood flow with electromagnetic flow probes remains the best available means of assessing individual vessel flow. Although the technique has found widespread application in experiments using large animals, its use in clinical situations has been limited by the relatively extensive vascular dissection required for placement of the probes and by the overestimation of true hepatic tissue blood flow, which occurs in the presence of portosystemic shunts. In addition, relative movements of the probes can give rise to errors and the need for repetitive calibrations. Total LBF in anesthetized subjects was determined to be approximately 1 L/min, of which about 25% was supplied by the hepatic artery (Schenk et al, 1962). Electromagnetic probes have been used intraoperatively to assess the hemodynamic status of the liver in cirrhosis (Ohnishi et al, 1987) after liver surgery or transplantation (Takaoka et al, 1990). A typical experimental preparation using electromagnetic flow probes is illustrated in Fig. 4.2.
Doppler Ultrasound
Invasive
Because the ultrasound probes are fitted directly to the vessel, such systems have been used successfully in the intraoperative (Henderson et al, 1991) and postoperative measurement of portal venous and hepatic arterial blood flow in liver transplant patients (Payen et al, 1990).
Noninvasive
In experiments performed on anesthetized dogs, good correlation was found between portal venous flow measured by a transcutaneous Doppler duplex system and electromagnetic flow probes fitted to the portal vein (Dauzat & Layrargues, 1989). However, the clinical application of transcutaneous Doppler ultrasound to obtain quantitative hepatic blood flow measurements in the portal vein have been more difficult (Burns et al, 1987). This technique has been utilized to assess postoperative hemodynamic consequences of portosystemic shunt procedures, such as transjugular intrahepatic portosystemic stent shunt (TIPS) (see Chapter 96; Fung et al, 1998), and for diagnosis of cirrhosis and portal hypertension (Iwao et al, 1997). In normal subjects, portal blood flow of 600 to 900 mL/min was reported (Brown et al, 1989; Moriyasu et al, 1986), whereas blood flow in the common hepatic artery was reported to be approximately 250 mL/min (Nakamura et al, 1989). The use of color Doppler has improved the suitability of the Doppler method for routine clinical use (Rosemurgy et al, 1997). The availability of ultrasound echo-enhancing materials, such as the galactose-based microbubble agent Levovist (Ernst et al, 1996), may further improve the precision of hepatic blood flow measurement and imaging using the Doppler technique.
Total Liver Blood Flow
Clearance Techniques
The rate of disappearance from the bloodstream of an indicator substance exclusively cleared by the liver is proportional to LBF. First applied to humans by Bradley and colleagues in 1945, indirect clearance methods of LBF measurement rely on the Fick equation. The flow measurement obtained by Bradley’s group depended on the fact that intravenously injected bromosulfophthalein is removed from the bloodstream and into the bile entirely by hepatocytes. They derived a value for the rate of hepatic bromosulfophthalein removal indirectly by determining the rate of intravenous infusion of dye that maintained the arterial concentration at a constant level, and by measuring the arteriovenous concentration difference of bromosulfophthalein, they were able to calculate total hepatic blood flow. The mean value obtained in a group of normal subjects was 1.5 L/min.
Other substances dependent on hepatocyte extraction into bile have also been applied, such as indocyanine green (Caesar et al, 1961). None of these substances achieve complete hepatic removal, and hepatic vein cannulation is necessary to allow calculation of the true extraction efficiency. Many investigators now use a simplified version of the original method, in which indocyanine green is administered as a bolus, instead of as an infusion, and hepatic extraction efficiency is determined from an analysis of the clearance curve derived from blood samples taken from a peripheral vein, instead of from the hepatic vein (Grainger et al, 1983). Limitations of this technique include extrahepatic shunting, which is common in liver disease and results in an underestimation of blood flow. Other hepatic clearance techniques have been used, such as colloidal clearance by the hepatic Kupffer cells (Dobson & Jones, 1952) and hepatocyte removal of galactose (Keiding, 1988), sorbitol (Zeeh et al, 1988), rose bengal (Combes, 1960), or propranolol (George, 1979). The more complete hepatic extraction of these substances overcomes the need to cannulate a hepatic vein in patients with normal liver function.
A modification of the colloid extraction method developed in the 1980s allows the derivation of the ratio of hepatic arterial to total LBF, termed the hepatic perfusion index. The basis of the technique is the ability to determine by dynamic scintigraphy the temporal separation of accumulating hepatic activity from the arterial and portal supplies after the intravenous administration of a bolus of technetium-99m–sulfur colloid (Fleming et al, 1981; Parkin et al, 1983).
Indicator Dilution
The indicator dilution method relies on the application of the Stewart-Hamilton principle, also derived from the Fick equation (Stewart, 1897). In principle, the hepatic blood flow is proportional to the amount of hepatic blood that has diluted an introduced indicator. This method involves the injection into the hepatic artery and portal vein of a labeled substance that is not removed by the liver; changes in hepatic vein concentration are measured by blood sampling or by monitoring the hepatic isotope activity with an external detector. Such a method is therefore independent of hepatocellular function and reliable, provided the indicator remains in the vascular space and is not excreted prior to sampling. In addition, sampling must not be done in such a manner that affects hepatic circulation, or not more than 30 mL/min in dogs (Cohn et al, 1969).
Portal vein flow may be determined separately using a modification of this technique by sampling portal blood after splenic vein or superior mesenteric artery injection (Chiandussi et al, 1968; Huet et al, 1973). Hepatic artery flow may be calculated as the difference between total hepatic flow and portal flow. In addition, a modified thermal dilution technique has been used to measure portal blood flow in humans (Biber et al, 1983). Indicator dilution methods overestimate true blood flow to hepatic tissue when intrahepatic or extrahepatic shunts are present, although it is possible to measure azygos blood flow by thermal dilution in patients with cirrhosis (Bosch & Groszmann, 1984).
Indicator Fractionation
The measurement of regional blood flow by fractional distribution of cardiac output was first described by Sapirstein in 1956. Briefly, a known amount of radioactive microspheres is injected into the left ventricle, and a reference sample is withdrawn from a peripheral artery at a known rate. The microspheres are then extracted from the various vascular beds, where they have lodged in proportion to the cardiac output. The hepatic arterial blood flow is determined directly by this method, but the portal flow contribution is found indirectly by addition of the flow values in the prehepatic splanchnic organs. Examination of the intrahepatic distribution of microspheres provides a means of assessing the pattern of arterial flow in different regions of the liver (Greenway & Oshiro, 1972). Because the microsphere method requires the postmortem removal of the organs of interest for radioactivity or colorimetric measurement, the additional determination of tissue weight enables flow per gram (i.e., tissue perfusion) to be calculated. Microspheres may be employed to determine the extent of portosystemic shunts: the fractional distribution in the liver may be measured with respect to systemic (lung) activity after portal vein injection, or it may be estimated by injecting a second radioactive microsphere directly into the splenic or mesenteric venous system (Groszmann et al, 1982).
Hepatic Tissue Perfusion
Inert Gas Clearance
By exploiting the fact that radioactive gases such as krypton (85Kr) and xenon (133Xe) distribute equally between tissue and blood according to a specific partition coefficient, the rate of clearance of such gases can be measured after their injection into the hepatic blood supply. After injection and rapid diffusion throughout the liver, the gas clears from the tissue into the blood and is almost completely eliminated from the body after a single passage through the lungs. The clearance rate is proportional to hepatic tissue perfusion, which may be calculated using a standard formula (Leiberman et al, 1978). Beta-emissions of 85Kr are recorded by a Geiger-Müller tube or semiconductor (silicon) detector placed on or immediately above the exposed liver surface, while the γ-emissions of 133Xe are monitored transcutaneously by a single scintillation crystal or a γ-camera; the latter device allows simultaneous measurement of hepatic tissue perfusion in many regions of interest.
Inert gas techniques involve minimal trauma to the patient, and their accuracy is not markedly affected by the presence of hepatic cellular disease or nonperfusion shunts; however, some variability does occur, even within the same subject, when multiple studies are performed. The first to use the inert gas method in the hepatic circulation were Aronsen and colleagues (1966), who recorded the γ-emissions of 133Xe after the injection of a saline solution of the isotope into the portal vein.
Laser Doppler Flowmetry
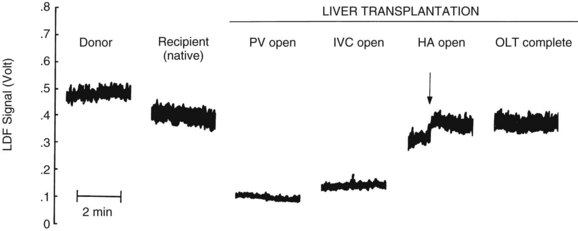
Laser Doppler flowmetry (LDF) is a more recent but established technique for the real-time measurement of microvascular red blood cell perfusion in the liver. By illuminating the tissue with low-power laser light and capturing the backscattered light with independent photodetectors, the Doppler shift of moving cells can be transmitted as an electrical signal. Linearity of the LDF signal from the liver with total organ perfusion has been shown (Almond & Wheatley, 1992; Shepherd et al, 1983, 1987), and the technique has been shown to be sensitive to rapid changes in organ flow (Almond & Wheatley, 1992). In a rat model, the technique provided a good measure of hepatic perfusion in vivo (Wheatley et al, 1993a; Fig. 4.3). LDF has also been used successfully to assess LBF during shock and resuscitation (Wang et al, 1995) and during drug-induced changes in LBF in the rat (Kurihara et al, 1992), and the technique has been applied successfully to measure LBF during liver transplantation in humans (Seifalian et al, 1997). A major drawback of the technique is that, owing to the small volume of tissue interrogated by the laser, the LDF signal can only be used to measure arbitrary, instead of absolute, blood perfusion in a single area. The development of laser Doppler perfusion imaging devices that estimate tissue perfusion by scanning an area of tissue using a laser may overcome the problem of signal calibration (Wardell et al, 1993).
In Vivo Fluorescent Microscopy
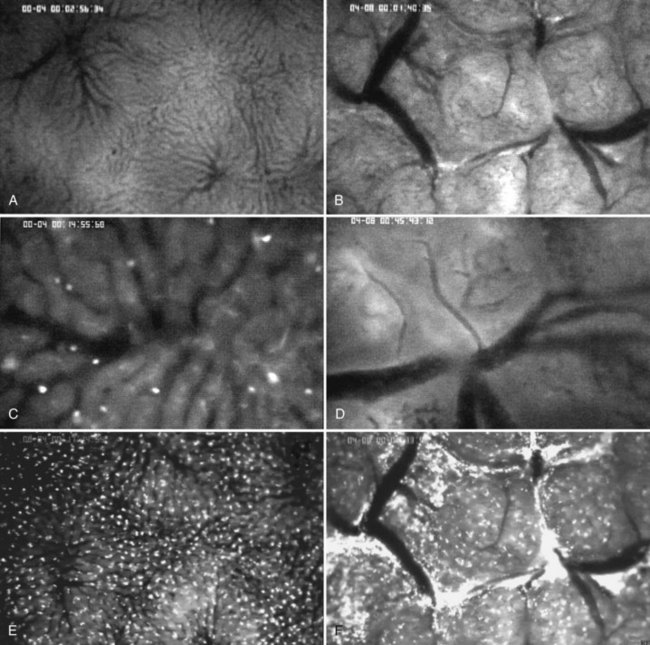
Intravital microscopy was first described in the microvessels of the frog tongue by Waller in 1846. Using this technique, individual sinusoids and terminal venules can be visualized, and changes in their diameters and the velocities with which erythrocytes pass through them can be seen (Menger & Messmer, 1991; Fig. 4.4). The introduction of fluorescent dyes has broadened the spectrum of in vivo microscopy in the liver from morphologic analysis to the study of pathologic events. However, from a hemodynamic point of view, intravital fluorescent microscopy has problems of interpretation (Sherman et al, 1990). In perfused liver, a 2.5-fold increase in portal venous blood flow has been found to be associated with only a 22% increase in sinusoidal red blood cell velocity, suggesting that changes in portal venous blood flow have only a minor effect on the capillary transit time (Sherman et al, 1996). In the regenerating liver; however, a 50% increase in portal venous blood flow resulted in a similar (66%) increase in sinusoidal red blood cell velocity (Wheatley et al, 1996; Zhang et al, 1997).
Near-Infrared Spectroscopy
Near-infrared spectroscopy is a new noninvasive technique that utilizes light transmission and absorption to measure hemoglobin and mitochondrial oxygenation. In contrast to visible light, which can only penetrate a few millimeters, near-infrared light (700 to 1000 nm) can be detected through up to 80 mm of tissue. The application of this technology to monitor liver oxygenation has been validated in models of endotoxic shock in pigs (Nadhum, 2006) and by intraoperative quantification of congestion and mitochondrial redox during hepatic vein occlusion in living-donor transplantation (Ohdan, 2003).
New and Future Developments
Orthogonal polarized spectral imaging has been used to obtain images of the liver microcirculation comparable to those of fluorescence microscopy but without the need for fluorescent dyes (Langer, 2001). This system relies on the absorbance of hemoglobin and can discriminate a single red blood cell in an individual capillary with a diameter of 5 microns. Studies with hand-held devices using this technology have been applied to healthy living donors to obtain sinusoidal red blood cell velocity and columetric blood flow (Puhl et al, 2003). A recent successor to spectral imaging is sidestream dark field imaging, whereby a light guide surrounded by diodes can detect both red and white blood cells in the microcirculation without the surface reflections (Czerny et al, 2009). Portal venous blood flow measured by noninvasive magnetic resonance imaging (MRI) has been found to correlate well with flow measured by Doppler ultrasound flow probes (Pelc et al, 1992); subsequently, the technique has been used to measure portal venous blood flow in human liver transplantation candidates (Kuo et al, 1995). Further methodology improvements in MRI (e.g., dynamic contrast enhancement) and in other techniques, such as positron emission tomography, may lead to their increasing use in the study of hepatic hemodynamics in humans (Chow et al, 2003).
Clinical Relevance
Hemorrhagic Shock, Hypoperfusion, and Ischemia-Reperfusion Injury
Total LBF decreases approximately in relation to the degree of the hemorrhage, and portal venous blood flow decreases in parallel to cardiac output; but similar to the coronary, pulmonary, and cerebral circulations, hepatic arterial flow does not decrease until extremely low blood pressures are reached. As a result, the hepatic oxygen supply tends to be maintained, although oxygen extraction greatly increases to preserve normal total oxygen consumption (Smith et al, 1979). Hepatic blood volume can increase significantly in cardiac failure and can compensate up to 25% of hemorrhage from its large capacitance vessels (Lautt, 2007).
The clinical entity known as shock liver
Related posts:
 Preoperative and postoperative nutrition in hepatobiliary surgery
Preoperative and postoperative nutrition in hepatobiliary surgery
 Cytokines in liver, biliary, and pancreatic disease
Cytokines in liver, biliary, and pancreatic disease
 Nonhepatic surgery in the cirrhotic patient
Nonhepatic surgery in the cirrhotic patient
 Liver transplantation for cholangiocarcinoma and other neoplastic diseases
Liver transplantation for cholangiocarcinoma and other neoplastic diseases
 Distal splenorenal shunt
Distal splenorenal shunt
 Interventional endoscopy: Technical aspects
Interventional endoscopy: Technical aspects
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


