CHAPTER 99 Intestinal Electrolyte Absorption and Secretion
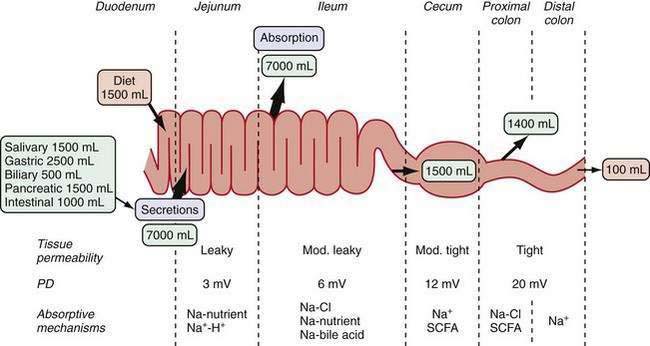
The gastrointestinal (GI) tract processes 8 to 9 L of fluid daily that is derived from oral intake and endogenous exocrine secretions. Intestinal fluid absorption is a process that functions with 98% efficiency, allowing only 100 to 200 mL to be excreted each day. The intestine also extracts nutrients, vitamins, and minerals; excludes destructive antigens and microbes; and excretes waste (Fig. 99-1). This multitasking is achieved by the unique tissue, cellular, and molecular architecture of the small and large intestine in combination with a complex array of intricate regulatory mechanisms (Fig. 99-2). Regulation is accomplished by cross-talk between endocrine and paracrine hormones, neurotransmitters, immunomodulators, and luminal factors. Remarkably, this orchestration proceeds smoothly on a daily basis; however, when the balance is perturbed, as occurs with an enteric infection, diarrhea ensues.
INTESTINAL ARCHITECTURE AND TRANSPORT
The structural and functional design of the intestine is optimally geared to its functions of absorbing nutrients and transporting fluids. In the small intestine, a 600-fold amplification of the absorptive surface is achieved by structural features, such as the circular folds of Kerckring (plicae circulares), villus-crypt architecture, and microvilli. Using a cylinder as the model, it has been estimated that the surface area of the small intestine is about 3300 cm2; the plicae circulare, villi, and microvilli amplify the surface area by factors of 3, 10, and 20, respectively, ultimately giving a surface area of about 2,000,000 cm2. In the large intestine, the spatial separation of crypts and surface cells allows efficient reabsorption of fluid. The overall architecture of the intestinal musculature can influence bulk fluid flow and transit time via changes in motility patterns (see Chapters 97 and 98), but the work of fluid transport occurs in the epithelia.
BASIC EPITHELIAL CELL MODEL
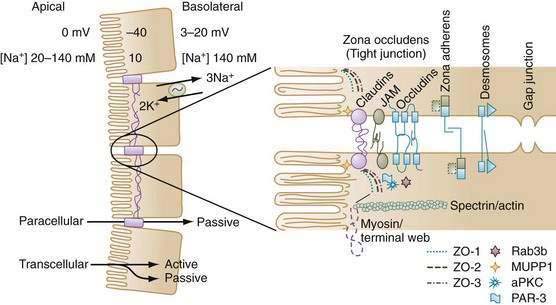
This basic cell model is modified by insertion of transporters into either the apical or basolateral membrane or by the characteristics of tight junctions that determine the unique qualities of a specific epithelial segment. A complex interaction of protein-sorting signals, cytoskeletal elements, and intracellular trafficking processes determines whether a newly synthesized protein is targeted to either the apical or basolateral membrane. For example, proteins with a glycosyl phosphatidyl inositol (GPI) anchor (e.g., alkaline phosphatase or carcinoembryonic antigen) are often associated with lipid rafts, and the GPI anchor serves to direct them toward the apical membrane.1 Membrane proteins destined to be delivered to the basolateral membrane carry specific membrane-sorting signals (amino acid sequences) in their cytoplasmic tails. In contrast, other proteins can insert randomly into either apical or basolateral domain, but they may be retained in the basolateral pole by specific components such as ankyrin.2
Regulation of intracellular trafficking ensures delivery of the right protein to the right membrane and is critical for establishing epithelial polarization and vectorial transport. When tight junctions are disrupted in vitro, diffusion and intermingling of apical and basolateral proteins in the fluid phase of the membrane result in a loss of epithelial cell polarity. There is some evidence that the distribution of Na+ pumps is altered during postischemic injury.3
The most prominent feature of epithelial cell polarity is targeting of the Na+,K+-ATPase pump to the basolateral membrane, for which expression of the beta subunit of Na+,K+-ATPase is critical. The Na+ pump is electrogenic, extruding three Na+ ions in exchange for two K+ ions, and thereby maintaining relatively low intracellular Na+ and high intracellular K+ concentrations compared with concentrations of these electrolytes in the extracellular environment (see Fig. 99-2). There also is greater membrane permeability for K+ over Na+, which favors diffusional exit of K+ from the cell over diffusional cellular entry of Na+. These features, in combination with the large number of intracellular proteins with fixed negative charges, lead to the characteristic negative intracellular potential difference compared with either the mucosal or serosal compartments.* Low [Na+] and electronegativity establish a favorable electrochemical gradient for passive Na+ entry into the cell. Functionally, the epithelial cell uses the energy of the favorable Na+ gradient to transport not only Na+ ions but also a variety of nutrients, vitamins, and electrolytes.
These properties provide the basic mechanisms of ion and water transport that apply to all epithelia. In the intestine, differences in transport can be seen along its cephalocaudal length as well as along the surface-crypt axis within a particular segment of intestine. Tissue- and segment-specific nuances arise from structural-functional and regulatory differences of both intracellular and intercellular proteins.4
SEGMENTAL HETEROGENEITY OF TRANSPORT
All intestinal segments from the duodenum to the distal colon have mechanisms for absorbing and secreting water and electrolytes. The diverse physiologic functions along the length of the GI tract are supported by the varied array of transporters encountered in its different segments. For example, the glucose- and amino acid-coupled transporters in the jejunum are well suited for absorption of large volumes of nutrients and water. The cecum, proximal colon, and distal colon, however, exhibit distinctly different transporters, with electrogenic Na+ absorption in the distal colon accomplishing the necessary final fluid extraction in preparation of feces.5–8 Different transporter molecules have been identified in specific segments of the GI tract. What is not clear, however, is why an individual transporter is located only in a specific segment of the intestine. For example, the DRA (down-regulated in adenoma) protein is an anion exchanger, and although anion exchange function is recognized in different segments of the intestine, DRA is predominantly expressed in the colon9–10 (see the later discussion of bicarbonate transport).
There also is segmental heterogeneity along the crypt-villus axis. Stem cells near the base of the crypt differentiate and migrate upward to form villus enterocytes in the small intestine or surface colonocytes in the large intestine while undergoing important changes in their transport and barrier properties (Fig. 99-3).11,12 As epithelial cells migrate away from the proliferative zone, the complexity of their tight junctions increases, the microvillus architecture of their apical membrane becomes more pronounced, and underlying cytoskeleton and signaling molecules undergo change; there also is increased expression of Na+ nutrient-coupled transporters, apical Na+-H+ exchangers, and brush border membrane hydrolases. In contrast, the levels of the Na+ pumps remain relatively constant and others, such as the signaling molecule adenylate cyclase and the cyclic adenosine monophosphate (cAMP)-associated Cl− channel CFTR (cystic fibrosis transmembrane conductance regulator), decrease in more mature villus cells.
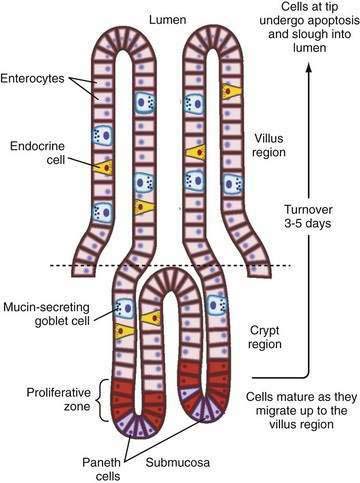
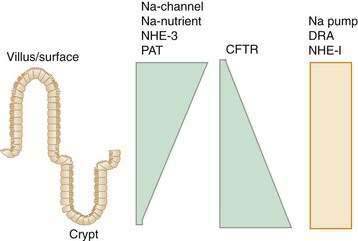
This spatial distribution of transporters (Fig. 99-4) is consistent with a model in which secretory function resides primarily in the crypts and absorption occurs in villus or surface cells. This dichotomy between absorptive surface cells and secretory crypt cells, however, is far from absolute; for example, in the colon, crypts absorb Na+ and fluid, and surface cells secrete Cl−.13,14 Thus, depending on their relative position along the crypt-villus axis, the crosstalk between transporters and their signaling molecules can vary and fine tune intestinal function. This segregation of absorptive and secretory functions might explain why, in diseases that selectively damage villi or surface epithelia—such as enteric infection, inflammatory bowel disease (IBD), and celiac disease—secretion predominates.
TIGHT AND LEAKY EPITHELIA
The paracellular space and junctional complexes between cells define the barrier function of epithelia. Epithelia with a low transepithelial voltage and low resistance are considered leaky, and those that exhibit a high transepithelial voltage and high resistance are considered tight. The tight junctions in villi have higher resistance than do those in crypts. Transepithelial resistance increases in a cephalocaudal direction (see Fig. 99-1).13
Since the 1990s, the model of paracellular transport and tight junctions has rapidly evolved from a static rigid barrier to a dynamic complex structure that is finely regulated (see Fig. 99-2). Movement through the space is exclusively passive, but it is influenced by electrical conductivity, charge selectivity, and its ability to be regulated. Cell-to-cell communications along the paracellular pathway occur in several discrete structures: zona occludens (ZO; tight junction), zona adherens (ZA), desmosomes, and gap junctions. The ZO is composed of several families of proteins that determine its physical and biological properties. For example, claudins belong to a family of 24 membrane-spanning proteins (24-27 kd) that form pores by interactions of the extracellular domains of claudins of adjoining cells; homotypic adhesion claudins are important in determining the charge selectivity of the tight junction.15,16
Additional proteins in the tight junction include occludins, junctional adhesion molecules (JAMs), and scaffolding proteins such as the zona occludens proteins (ZO-1, ZO-2) and multi-PDZ domain protein 1 (MUPP-1). The scaffolding proteins serve to link membrane proteins to an array of protein kinases, phosphatases and, via filamentous actin, to myosin in the terminal web, thereby influencing paracellular permeability.17,18 For example, disruption of tight junctions by enteropathogenic Escherichia coli is specifically associated with protein kinase Cζ activation.19 Another junctional complex that allows cell-to-cell interaction is the zona adherens. In epithelia, the zona adherens primarily is made up of E-cadherins, 120-kd transmembrane glycoproteins, with extracellular motifs that engage in calcium-dependent homotypic interaction with cadherins of adjoining cells. Intracellularly, cadherins bind to a family of adhesion molecules, the catenins, which in turn anchor to a dense actin-filament network. Alterations in cadherin-catenin distribution or function have been implicated in carcinogenesis.20
Desmosomes are junctional complexes that are structurally similar to zona adherens junctions, although instead of actin, they link to intermediate filaments through a dense plaque of intracellular anchor proteins. Gap junctions have a unique function: They bridge gaps between cells, thus allowing neighboring cells to exchange small molecules. They are made up of an assembly of connexins, a four-pass membrane-spanning protein, six of which join to form a hemichannel. When these hemichannels in two adjoining cells are aligned, they form a continuous pore that connects the interior of the two cells.1
TRANSEPITHELIAL TRANSPORT
Our current understanding of the movement of ions, solutes, and fluid across epithelia is gleaned from a combination of in vitro studies using reductionist models of cell lines or isolated epithelial sheets, and from complex in vivo methodologies such as the triple-lumen perfusion technique. All these models underscore that transepithelial ion (largely Na+) movement from the mucosa to the serosa drives fluid absorption, whereas net ion (largely Cl−) movement in the reverse direction drives fluid secretion. Although different approaches help elucidate a complex mechanism, at times they give confounding results. For example, some in vitro studies report decreased Cl− secretion and increased Na+ absorption in the jejunum of cystic fibrosis patients, implying that the intestinal manifestations of the disease are due to hyperabsorption of water. In contrast, in vivo studies show decreases in both Cl− secretion and passive Cl− absorption, suggesting that rather than a hyperabsorption of fluid, the severity of the disease is reflected by decreased fluid absorption.21
WATER MOVEMENT
Although water movement is a major property of the intestine, the mechanism(s) of intestinal water transport have not been clearly delineated. The movement of water is inextricably linked to the movement of solutes, in response to osmotic gradients. The standing-gradient hypothesis of water absorption suggests that even a small increase (2-3 mOsm) in the osmolarity of the intercellular and subepithelial spaces can cause movement of water across the epithelium, both through and around the cells.22 In the early 1950s, “water pores” were postulated to explain transepithelial water movement, but it was not until the remarkable discovery of the aquaporin (AQP) family of water transporters that a role of specific membrane proteins was implicated in erythrocyte and renal water transport.23,24 Although AQP2, AQP3, and AQP7 have been localized to the small intestine and AQP1, AQP3, AQP4, and AQP8 have been localized to the large intestine, the specific intestinal apical water channel, if any, has eluded discovery.25 Wright and associates proposed that the apical Na+-glucose transporter (SGLT) also may be able to transport water, perhaps as much as 5 L/day,26,27 but whether this can compensate for the puzzling lack of functional apical AQPs remains to be determined.
CHANNELS, CARRIERS, AND PUMPS
Small hydrophobic and uncharged molecules move across the lipid bilayer of the cell by diffusion, the rate of transport determined by the concentration gradients and diffusion coefficients (Fig. 99-5). Oxygen, carbon dioxide, fat-soluble vitamins, and unconjugated bile acids are examples of substances transported by diffusion. Because the majority of ions and solutes cannot cross the phospholipid membrane by diffusion, the cell employs an array of distinct integral membrane proteins, including channels, carriers, and pumps to cross cell membranes (Fig. 99-5).1
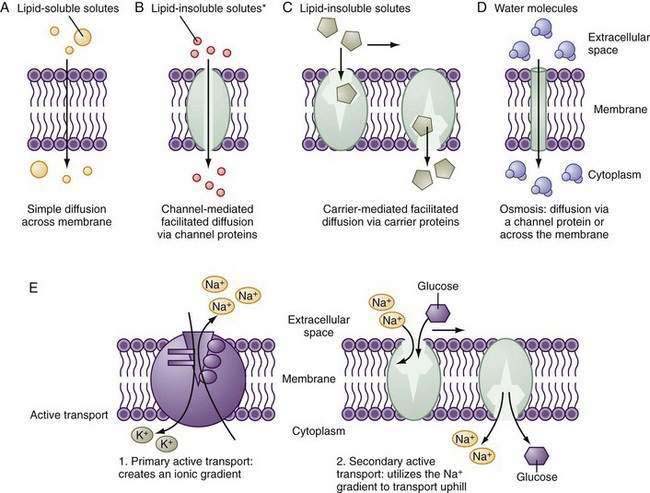
Channels are pores that allow the swift (>106 ions/sec) and controlled (by rapid opening and closing) transit of ions across the membrane, driven by the electrochemical gradient. The advent of molecular cloning techniques, patch clamp methodology (which allows the measurement of function of single channels), and membrane protein crystallography has greatly advanced our knowledge of how these proteins function. Channels tend to be ion selective. For example, Na+ channels exclude K+ despite its same charge and smaller size. Selectivity is determined by the hydration radius of the ion and the physiochemical nature of the pore. The overall transport of a particular ion is determined by the electrochemical gradient, the density of channels, and the gating (open-close time) of the channel; gating may be modulated by voltage, ion, concentration, or intracellular regulation. Mutations of critical residues in the channel protein can have dire functional consequences; for example, in cystic fibrosis, specific mutations of the CFTR affect the ability to transport chloride and bicarbonate.28,29
ION TRANSPORTERS
APICAL SODIUM CHANNEL
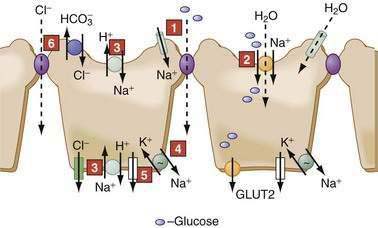
In the GI tract, the surface epithelial cells of the distal colon and rectum exhibit electrogenic Na+ absorption against a fairly steep concentration gradient. The downhill electrochemical gradient created by the Na+ pump drives Na+ entry via an apical membrane Na+-specific ion channel (Fig. 99-6), which belongs to the family of epithelial Na+ channels (ENaCs). Members of the ENaC family are found in many epithelia.30–32 They are multimeric proteins composed of α, β, and γ subunits; they exhibit a high sensitivity to the diuretic amiloride; and they are stimulated by mineralocorticoids and cAMP. Colonic ENaCs are inhibited by increases in intracellular Ca2+. Unlike many other channels that regulate transport by gating, ENaCs modulate transport by varying the channel density in the cell membrane; this variation may be accomplished through changes (increases or decreases) in synthesis (e.g., aldosterone) or exocytosis (e.g., cAMP, vasopressin) of the channels. Additionally, both aldosterone and cAMP block the association of ENaC with Nedd4-2, a ubiquitin protein ligase, which normally flags the protein for degradation; this block of degradation increases ENaC. Mutations in this pathway can result in the increased Na+ absorption and hypertension characteristic of Liddle’s syndrome, an autosomal dominant disorder with features of hyperaldosteronism that is a cause of infantile hypertension.
NUTRIENT-COUPLED SODIUM TRANSPORT
Glucose transport processes, elegantly elucidated by Wright and coworkers, provide a good example of nutrient transport.33 Transport across SGLT1 is electrogenic (2 Na+ to 1 glucose), is stereospecific (d-isomer), and transports galactose but not fructose.33 Glucose exit across the basolateral membrane occurs via a separate family of facilitated diffusion carriers, the glucose transporters (GLUT-2) (see Fig. 99-6). Fructose enters the cell via another member of this family, GLUT-5, and exits via GLUT-2.
Although it is clear that Na+ and glucose absorption stimulates water absorption, the mechanism is not fully delineated. The classic explanation is that basolateral exit of glucose creates a hypertonic compartment in the paracellular space, thereby generating an osmotic gradient for the entry of fluid from the lumen. Some enticing evidence suggests that secondary to transcellular transport through SGLT, passive processes triggers the contraction of the actomyosin ring in the terminal web, resulting in increased paracellular permeability to glucose and to water. Additionally, SGLT activation results in a protein kinase-dependent recruitment of GLUT2 to the apical membrane, which then serves as a high-capacity, low-affinity route for sugar entry during feeding.34,35 Evidence suggests that SGLT can serve as a water channel (210 water to 2 Na+ to 1 glucose) and might account for up to 5 L of fluid reabsorption in the fed state.36 How molecular regulation of transporters is translated into net nutrient absorption during feeding in vivo is a critical area of inquiry. For a description of similar advances made in our understanding of amino acid and vitamin transport, see Chapters 100 and 101.
SODIUM-HYDROGEN EXCHANGERS
Ten mammalian isoforms of NHE have been cloned. NHE1-4 and 6-9 exhibit species- and segment-specific distribution in the GI tract.37–39 NHE1 is a plasma membrane protein found in epithelial and nonepithelial cells. It is expressed on epithelial basolateral membranes and functions as the housekeeper regulator of intracellular pH, cell volume, and growth. NHE2 and NHE3 are apical membrane proteins restricted to epithelia and are the major conduits for electroneutral Na+ absorption in the intestine (see Fig. 99-6). NHE2 is expressed throughout the GI tract, but maximal expression is in the proximal colon. NHE3 is considered a marker for the absorptive cells of the small intestine and colon; it is expressed only in the villus or surface cells, and not in the crypts. NHE4 is located in the basolateral membrane and is primarily expressed in gastric parietal and chief cells, where it might have a role in acid secretion. NHE5 and NHE10 are not expressed in the GI tract, and the roles of NHE6-9 remain to be determined.
NHE activity is differentially modulated by neural, paracrine, or endocrine stimuli through intricate scaffolding complexes that include the exchanger itself, a family of NHE regulatory factors (NHERFs) that act as a bridge between the exchanger, and a variety of kinases and phosphatases.40,41 Different stimuli use differing scaffolding complexes to exert their effect. For example, glucocorticoids stimulate Na+ absorption and up-regulate NHE3 but not NHE1, 2, or 4, consistent with their respective roles in vectorial transport and housekeeping. Glucocorticoids act via a serum and glucocorticoid inducible kinase, SGK1; SGK1 stimulates the activity of NHE3 by interacting directly with NHERF2.
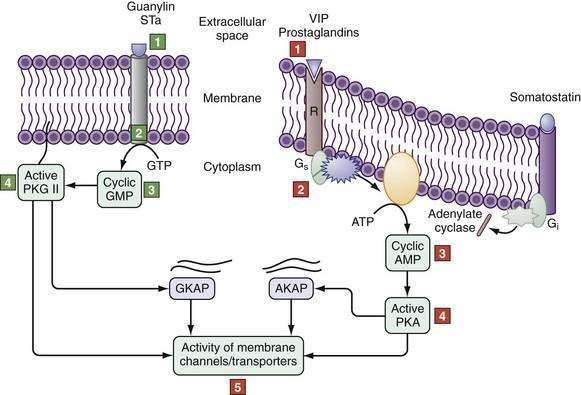
Alternatively, cAMP inhibits NHE3 by activating protein kinase A, which is recruited to the C-terminus of NHE3 by NHERF1, NHERF2, and an additional cytoskeletal protein, ezrin. In this location, PKA induces its inhibitory effect by phosphorylating NHE3. Cyclic guanosine monophosphate (cGMP) can inhibit NHE3 by triggering the formation of a complex between cGMP-dependent protein kinase II (cGKII) and NHERF2, which anchors cGMP kinase protein (GKAP). Activation of guanylate cyclase C by guanylin or E. coli–stable toxin A increases cGMP content near the brush border to locally activate cGKII (Fig. 99-7), which then inhibits NHE3 activity.40
CHLORIDE (ANION) ABSORPTION
The transepithelial, lumen negative potential difference contributes to the passive movement of Cl− and other anions via the paracellular pathway in the jejunum.7 While coupled Cl−-HCO3− and Na-H exchangers govern electroneutral transport in the ileum and proximal colon, sodium-independent Cl−-HCO3− exchange occurs in the distal colon (see Fig. 99-6) (see the discussion of HCO3− transport, later).
CHLORIDE SECRETION
The principal driving force for the secretion of fluid is the transcellular movement of Cl− from the serosal to the luminal compartment. Na+ and water follow passively in response to the ensuing electrical and osmotic gradients (Fig. 99-8). The small and large intestine exhibit a basal rate of Cl− secretion that is maintained by the interplay of cell volume, [Cl]i, and paracrine, autocrine, neuronal, endocrine, luminal, and immune modulators. Disruptions in the balance of these regulatory processes can lead to secretory diarrhea.
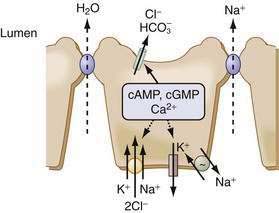
Several epithelia in the GI tract exhibit electrogenic Cl− secretion. Although there are some tissue-specific regulatory differences, the mechanisms underlying this secretion are remarkably similar. The Na+ pump provides the driving force, Cl− enters the cell across the basolateral membrane via an electroneutral cotransporter (NKCC1) that couples the movement of 1Na+:1K+:2Cl−, and Cl− leaves the cell via specific channels on the apical membrane. The Na+ entering the cell via NKCC1 exits via the Na+ pump, and the K+ leaves via K+ channels either on the apical or the basolateral membrane. This complex interplay of transporters is an elegant demonstration of cellular economy. The NKCC1 cotransporter effectively moves 2 Cl− and 1 K+ uphill for the expenditure of a single Na+ ion. The pump-to-leak relation between K+ channels and the Na+ pump helps to maintain the interior of the cell as electronegative, thereby providing the driving force for Cl− exit. Basolateral K+ exit electrically balances the large Cl− flux across the apical membrane. NKCC cotransporters belong to a superfamily of cation transporters and are characterized by their inhibition by the loop diuretics bumetanide and furosemide.42–44
Related posts:





Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


