CHAPTER 42 Esophageal Neuromuscular Function and Motility Disorders
ESOPHAGEAL MOTOR AND SENSORY FUNCTION
OROPHARYNX AND UPPER ESOPHAGEAL SPHINCTER
The oral cavity and pharynx are critically involved in the task of ingesting food. Within the oral cavity, the lips, teeth, hard palate, soft palate, mandible, floor of the mouth, and tongue serve functions in masticating, containing, and forming food into a bolus suitable for transfer to the pharynx. The pharynx is a hollow cavity separated into three segments (Fig. 42-1): nasopharynx, oropharynx (oral pharynx), and hypopharynx (laryngeal pharynx). The nasopharynx extends from the base of the skull to the distal edge of the soft palate. Although not part of the alimentary tract, muscles in the nasopharynx contribute to elevating the soft palate and sealing the nasopharynx during swallowing, preventing nasopharyngeal regurgitation. The oropharynx extends from the soft palate to the base of the tongue. The inferior margin of the oropharynx is demarcated by the valleculae anteriorly and the mobile tip of the epiglottis posteriorly. The hypopharynx extends from the valleculae to the inferior margin of the cricoid cartilage and includes the upper esophageal sphincter (UES).

The intrinsic muscles of the pharynx are the superior, middle, and inferior pharyngeal constrictors (see Fig. 42-1). The constrictors overlap and insert into a collagenous sheet, the buccopharyngeal aponeurosis. The superior pharyngeal constrictor arises from the pterygoid hamulus, pterygomandibular raphe, mandible, and tongue; passes posteromedially; and inserts to the posterior raphe. The middle constrictor arises from the hyoid bone and stylohyoid ligament, passes posteromedially, and inserts in the posterior median raphe. The inferior constrictor is composed of the thyropharyngeus (superior part) and the cricopharyngeus (inferior part). The thyropharyngeus arises from the thyroid cartilage, passes posteromedially, and inserts in the median raphe. The cricopharyngeus has superior and inferior components, each of which arise bilaterally from the sides of the cricoid lamina; the superior fibers course posteromedially to the median raphe and the inferior fibers loop around the esophageal inlet without a median raphe. Killian’s triangle, a triangular area of thin muscular wall is formed posteriorly between these components of the cricopharyngeus and is the most common site of origin for pharyngeal pulsion diverticuli.
The pharynx also contains five single or paired cartilages: the epiglottic, arytenoid, cuneiform, corniculate, and cricoid (see Fig. 42-1). The spaces formed between the lateral insertion of the inferior constrictor and the lateral walls of the thyroid cartilage are the pyriform sinuses that end inferiorly at the cricopharyngeus muscle, separating the pharynx from the esophagus. The larynx and trachea are suspended in the neck between the hyoid bone superiorly and the sternum inferiorly. A number of muscles, categorized as the laryngeal strap muscles, contribute to this suspension and, together with the intrinsic elasticity of the trachea, permit the larynx to be raised and lowered. The hyoid bone also serves as the base for the tongue that rests on it. Laryngeal movement is crucial to the successful enactment of the swallow response because the laryngeal inlet is closed and physically removed from the bolus path in the course of a swallow. Failure to achieve this synchronized laryngeal movement can result in aspiration.
The pharyngeal muscles are densely innervated with motor fibers coming from nuclei of the trigeminal, facial, glossopharyngeal, and hypoglossal nuclei as well as nucleus ambiguus and spinal segments C1 to C3. The innervation of the major pharyngeal muscles is as follows: mylohyoid, tensor veli palatini, and anterior digastric muscles (trigeminal nerve); stylohyoid and posterior part of the digastric (facial nerve); stylopharyngeus (glossopharyngeal nerve); levator veli palatini, palatopharyngeus, salpingopharyngeus, thryroarytenoid, arytenoid, pharyngeal constrictors, and cricopharyngeus (vagus nerve); thyrohyoid, geniohyoid, and tongue (hypoglossal nerve).1 Nucleus ambiguus is the vagal nucleus responsible for innervation of the striated muscle of the pharynx, larynx, and esophagus.2 All motor neurons within the nucleus ambiguus seem to participate in swallowing with those innervating the esophagus situated rostrally and those innervating the larynx more caudally.3
The muscular components of the UES are the cricopharyngeus, adjacent esophagus, and adjacent inferior constrictor (see Chapter 41, Fig. 41-3). The cricopharyngeus contributes the zone of maximal UES pressure, which is about 1 cm in length.4 The closed sphincter has a slit-like configuration with the cricoid lamina anterior and the cricopharyngeus making up the lateral and posterior walls. Not surprisingly, resting UES pressure is markedly asymmetrical with greatest values anteriorly and posteriorly.5 Neural input via vagal trunks originating in nucleus ambiguus to the UES is required for maintenance of high resting pressure and for the coordination of relaxation with swallowing.6 Cessation of motor neuron firing, or administration of curare, causes relaxation, whereas increased spike activity increases tone. Vagal transection abolishes contractile activity in the cricopharyngeus and inferior pharyngeal constrictor muscles.7
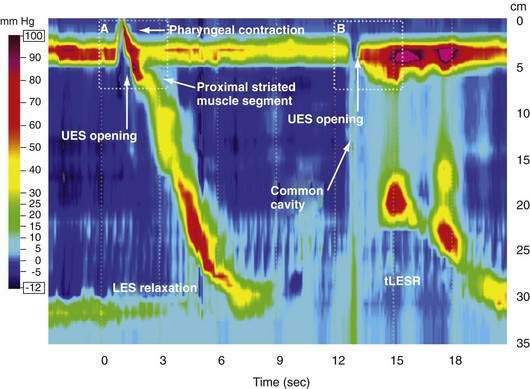
Manometric evaluation of UES function is difficult because it is a short, complex anatomic zone that moves briskly during swallowing. Furthermore, UES pressure measurement is heavily influenced by recording methodology due to its marked asymmetry and to the fact that pharyngeal stimulation by the measurement device stimulates sphincter contraction. Thus, it is not surprising that there is great variability in reported “normal” UES pressure ranges and it is currently impossible to define a meaningful normal range.8 UES relaxation during swallowing also poses substantial recording challenges, making for great variability in technique and interpretation among centers. However, high-resolution manometry using solid state technology has become clinically available, and this technology permits accurate tracking of UES relaxation and intrabolus pressure changes during swallowing (Fig. 42-2).
The main function of the UES is to maintain closure of the proximal end of the esophagus unless opening is required for either swallowing or belching.9 It constitutes an additional barrier to refluxed material entering the pharynx and prevents air from entering the esophagus by contracting in synchrony with inspiration. Inspiratory augmentation is most evident during periods of low UES pressure and can be exaggerated in individuals experiencing globus sensation.10 Balloon distention of the esophagus stimulates UES contraction with the effect being more pronounced with more proximal balloon positions.11 However, when the distension pattern of gas reflux is simulated using a cylindric bag or rapid air injection into the esophagus, UES relaxation rather than contraction occurs.4 Belch-induced UES relaxation is also associated with glottic closure.12 Stress augments UES pressure13 and anesthesia14 or sleep15 virtually eliminates it. Neither experimental acid perfusion of the esophagus16 nor spontaneous gastroesophageal acid reflux alters continuously recorded UES pressure in either normal volunteers15 or in individuals with peptic esophagitis.16
THE PHARYNGEAL SWALLOW
The oral phase of swallowing is largely voluntary and highly variable. Disorders of the oral phase of swallowing occur with many conditions characterized by global neurologic dysfunction such as head trauma, cerebral tumors, or chorea. Detailed discussion of these conditions can be found in texts on swallow evaluation and therapy.17,18 The pharyngeal swallow is the complex coordinated contraction that transfers oral contents into the esophagus. A typical individual swallows about 600 times a day without giving significant thought or effort to the activity.19 Afferent sensory fibers capable of triggering the pharyngeal swallow travel centrally via the internal branch of the superior laryngeal nerve (from the larynx) and via the glossopharyngeal nerve (from the pharynx).20 These sensory fibers converge before terminating in the medullary swallow center.21
Although understood physiologically as the patterned activation of motor neurons and their corresponding motor units, swallowing is clinically evaluated in mechanical terms and best evaluated by videofluoroscopic or cineradiographic analysis. The pharyngeal swallow rapidly reconfigures pharyngeal structures from a respiratory to an alimentary pathway and then reverses this reconfiguration within one second. The pharyngeal swallow response can be dissected into several closely coordinated actions: (1) nasopharyngeal closure by elevation and retraction of the soft palate, (2) upper esophageal sphincter (UES) opening, (3) laryngeal closure, (4) tongue loading (ramping), (5) tongue pulsion, and (6) pharyngeal clearance. Precise coordination of these actions is an obvious imperative and to some degree the relative timing of these events is affected either by volition or by the volume of the swallowed bolus (Fig. 42-3).22
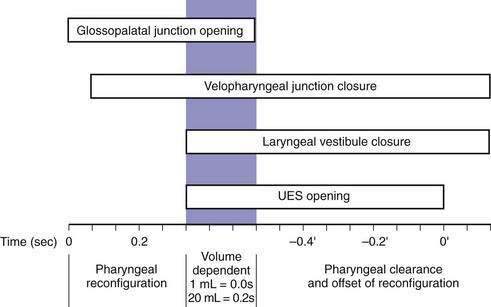
The most fundamental anatomic reconfiguration required to transform the oropharynx from a respiratory to a swallow pathway is to open the inlet to the esophagus and seal the inlet to the larynx. These events occur in close synchrony. UES opening occurs by laryngeal elevation and anterior traction via the hyoid axis.23,24 The mechanical determinants of laryngeal vestibule closure are laryngeal elevation and anterior tilting of the arytenoid cartilages against the base of the epiglottis.17 Thus, analyzing the efficacy of either of these events inevitably focuses on laryngeal elevation.25 UES relaxation occurs at roughly the same degree of laryngeal elevation regardless of swallow volume, and precedes sphincter opening by about 0.1 second. It is critical to recognize the distinction between UES relaxation and UES opening. UES relaxation occurs due to cessation of excitatory neural input while the larynx is elevating. Once the larynx is elevated, UES opening results from traction on the anterior sphincter wall caused by contraction of the suprahyoid and infrahyoid musculature that also results in a characteristic pattern of hyoid displacement.23,24
The two main determinants of bolus transport out of the oropharynx are the action of the tongue and of the pharyngeal constrictors. Tongue motion varies substantially with swallow conditions and propels most of the bolus into the esophagus prior to the onset of the pharyngeal contraction with larger volume swallows.26 On the other hand, the propagated pharyngeal contraction has similar propagation and vigor regardless of bolus volume.27 However, the propagated pharyngeal contraction is more involved with the process of clearance than of bolus propulsion; it strips the last residue from the pharyngeal walls. Upper esophageal sphincter closure coincides with the arrival of the propagated pharyngeal contraction as evident by the fixed time relationship between these events.24 However, the contractile activity of the sphincter has an added dimension as well, exhibiting increased electromyographic activity during laryngeal descent.28 The magnitude of this postdeglutitive contraction is further augmented by either sphincteric or proximal esophageal distention resulting in a grabbing effect such that the sphincter and laryngeal descent complement each other to clear residue from the hypopharynx.29 This clearing function probably acts to minimize the risk of post-swallow aspiration by preventing residual material from adhering to the laryngeal inlet when respiration resumes.
ESOPHAGUS
The esophagus is a 20- to 22-cm muscular tube with a wall composed of skeletal and smooth muscle (see Chapter 41, Fig. 41-1). The proportion of each muscle type is species dependent, but in man, the proximal 5% is striated, the middle 35% to 40% is mixed with an increasing proportion of smooth muscle distally, and the distal 50% to 60% is entirely smooth muscle.30 The bundles of the outer (longitudinal) muscle arise from the cricoid cartilage receiving slips from the cricopharyngeus and pass dorsolaterally to fuse posteriorly about 3 cm distal to the cricoid cartilage. This arrangement results in a posterior triangular area devoid of longitudinal muscle, Laimer’s triangle. Distal to Laimer’s triangle the longitudinal muscles form a continuous sheath of uniform thickness around the esophagus.31 The adjacent, inner muscle layer is formed of circular or, more precisely, helical muscle also forming a sheath of uniform thickness throughout the length of the esophageal body. The overlapping helices exhibit decreasing degree of helicity moving distally ranging from 60 degrees in the proximal esophagus to nearly 0 degrees in the most distal esophagus.32
The extrinsic innervation of the esophagus is via the vagus nerve (see Chapter 41). Fibers innervating the striated muscle are axons of lower motor neurons with cell bodies situated in nucleus ambiguus, whereas the smooth muscle esophagus is innervated by the dorsal motor nucleus of the vagus.33,34 Efferent nerve fibers reach the cervical esophagus by the pharyngoesophageal nerve35 and histologic studies show that vagal efferents synapse directly on striated muscle neuromuscular junctions.36,37 The vagus nerves also provide sensory innervation. In the cervical esophagus this is via the superior laryngeal nerve with cell bodies in the nodose ganglion, whereas in the remainder of the esophagus sensory fibers travel via the recurrent laryngeal nerve or, in the most distal esophagus, via the esophageal branches of the vagus. Histologic studies demonstrate many free nerve endings in the mucosa, submucosa, and muscular layers.2,38 Additionally, a few encapsulated structures resembling spindles have been described in humans. These vagal afferents are strongly stimulated by esophageal distention.
The esophagus also contains an autonomic nerve network, the myenteric plexus, located between the longitudinal and circular muscle layers.39 Myenteric plexus neurons are sparse in the proximal esophagus and their function in that region is unclear because the striated muscle is directly controlled by somatic motor fibers. On the other hand, the thoracic esophagus receives innervation from preganglionic neurons in the dorsal motor nucleus of the vagus that then synapse in myenteric plexus ganglia, relay neurons between the vagus and the smooth muscle. The ganglia of the myenteric plexus are more numerous in the distal esophagus than in the striated muscle region, but throughout, they are still far less dense and smaller than in other regions of the gut.40,41 A second nerve network, the submucosal or Meissner’s plexus, is situated between the muscularis mucosa and the circular muscle layer, but this is exceedingly sparse with few ganglia in the human esophagus.41
Properties of Esophageal Peristalsis
The esophagus does not normally exhibit spontaneous contractions and its intraluminal pressure closely reflects pleural pressure, becoming negative during inspiration. However, swallowing or focal distention initiates peristalsis. Primary peristalsis is that which is initiated by a swallow and traverses the entire length of the esophagus; secondary peristalsis can be elicited in response to focal esophageal distention with air, fluid, or a balloon, beginning at the point of distention.42 The mechanical correlate of peristalsis is a stripping wave that milks the esophagus clean from its proximal to distal end. The propagation of the stripping wave corresponds closely with that of the manometrically recorded contraction such that the point of the inverted “V” seen fluoroscopically at each esophageal locus occurs concomitantly with the upstroke of the pressure wave. The likelihood of achieving complete esophageal emptying from the distal esophagus is inversely related to peristaltic amplitude such that emptying becomes progressively impaired with peristaltic amplitudes of 30 mm Hg or less.43
Another essential feature of peristalsis is deglutitive inhibition. A second swallow, initiated while an earlier peristaltic contraction is still progressing in the proximal esophagus, causes rapid and complete inhibition of the contraction induced by the first swallow.44 If the first peristaltic contraction has reached the distal esophagus, it may proceed distally for a few seconds after the second swallow, but its amplitude then diminishes until it disappears.45 Deglutitive inhibition in the distal esophagus is attributable to hyperpolarization of the circular smooth muscle and is mediated via inhibitory ganglionic neurons in the myenteric plexus. Deglutitive inhibition can be demonstrated experimentally in the esophagus by creation of an artificial high-pressure zone with an intraluminal balloon.46 The artificial high-pressure zone is created by distending the esophageal lumen with a balloon and recording intraluminal pressure between the balloon and the esophageal wall. Once the high pressure zone is established in the normally flaccid tubular esophagus, deglutitive inhibition is evident by relaxation of the artificial high pressure zone commencing concurrently with the swallow.
The physiologic control mechanisms governing the striated and smooth esophageal musculature are distinct. The striated muscle of the esophagus receives exclusively excitatory vagal innervation and its peristaltic contraction results from sequential activation of motor units in a craniocaudal sequence. These fibers release acetylcholine and stimulate nicotinic cholinergic receptors on the motor endplates of the striated muscle cells. Physiologic evidence of this arrangement was provided by an ingenious series of experiments using the nerve suture technique.2 In these experiments, the vagal branch innervating the esophagus was severed and the central end anastomosed to the peripheral end of the also severed spinal accessory nerve. Thus, after a period of nerve regeneration, the vagal branch effectively innervates the sternocleidomastoid and trapezius muscles. Occurrence of excitatory vagal discharges can then be surmised from the contractile activity of these readily accessible muscles. Nerve suture experiments demonstrated several properties of vagal control of esophageal striated muscle: (1) vagal efferent fibers exhibit no spontaneous discharge but fire in spike bursts during primary or secondary peristalsis; (2) once activated, vagal fibers innervating different levels of the esophagus fire sequentially, demonstrating peristaltic programming by the medullary swallow center; (3) peristaltic vagal motor discharges are potentiated by stimulation of afferent fibers from the esophagus (designed to mimic the effect of a bolus being pushed ahead of the contraction); (4) peristaltic vagal motor discharges are stronger during primary than secondary peristalsis; and (5) vagal motor fibers are inhibited during the pharyngeal stage of deglutition or after distention of a proximal esophageal segment supporting the notion that deglutitive inhibition has a central origin. Thus, there is substantial evidence that peristalsis in the striated muscle esophagus is controlled by the swallowing center of the medulla in much the same way as is the oropharyngeal musculature.
The vagus nerves also exhibit control of primary peristalsis in the smooth muscle esophagus. Deviation of a swallowed bolus at the level of the cervical esophagus (thereby eliminating the potential for bolus-initiated afferent feedback) does not eliminate the primary peristaltic contraction in the distal esophagus.47 Furthermore, primary peristalsis of the smooth muscle persists even after curarization. Because curarization paralyzes the oropharyngeal and cervical esophagus, the persistence of distal peristalsis in these experiments strongly suggests that it is triggered by the medullary swallowing center that can elicit the entire motor sequence of primary peristalsis without receiving afferent feedback.
The mechanism of vagal control of the smooth muscle esophagus is more complex than that of the striated muscle because vagal fibers synapse on myenteric plexus neurons rather than directly on muscle cells. Experimentally, vagal stimulation either excites or inhibits esophageal musculature depending on the stimulation parameters used.48,49 In the esophagus of the opossum, vagal or swallow-induced stimulation causes depolarization with superimposed spikes on longitudinal muscle but an initial hyperpolarization followed by depolarization and spike burst on circular muscle.50 With swallowing initiated by superior laryngeal nerve stimulation, the response characteristics of single nerve fibers participating in smooth muscle peristalsis could be divided into two groups.51 Activity of short latency vagal fibers correlated temporally with the onset of the deglutitive inhibition while the activity of long latencies fibers was temporally correlated with the onset of contraction at each esophageal locus. Thus, activity of neurons in the dorsal motor nucleus of the vagus reflects several properties of primary peristalsis in the smooth muscle esophagus including deglutitive inhibition and both the speed and vigor of peristaltic contraction.
Control of peristalsis may also arise in the myenteric plexus. Stimulation of decentralized vagal efferents evokes peristalsis similar to that seen with swallowing and this is obliterated with transection across the smooth muscle esophagus, suggesting that an intact intramural neural myenteric plexus is necessary for peristaltic propagation. In contrast, transection across the striated muscle in the proximal esophagus does not inhibit peristaltic progression across the transection site or distally.47 Further evidence supporting the potential autonomy of peripheral mechanisms is that distention anywhere within the smooth muscle esophagus will elicit secondary peristalsis despite extrinsic denervation.52
Regardless of central or ganglionic control, esophageal smooth muscle contraction is ultimately elicited by ganglionic cholinergic neurons. Swallow-induced peristalsis is highly atropine sensitive, can be augmented by cholinergic agonists, and is inhibited by acetylcholinesterase.53,54 Less clear, however, are the control mechanisms for the direction and velocity of the peristaltic wavefront. Nerve conduction studies indicate that neural stimuli initiated by swallowing propagate with a speed of 5 to 6 m per second and therefore reach the ganglionic neurons along the length of the esophagus essentially simultaneously.51 However, the latency between the arrival of the vagal stimulus and muscle contraction progressively increases moving aborally. In humans, the latent period is two seconds in the proximal smooth muscle esophagus and five to seven seconds just proximal to the lower esophageal sphincter (LES). The in vitro correlate of this is that when electrically stimulated, distal esophageal muscle strips exhibit longer latencies to contraction than do strips from the proximal esophagus.55,56 The latency gradient can be changed by varying vagal stimulation parameters or by pharmacologic manipulation suggesting it to be the result of an interaction between the initial inhibition and subsequent excitation of esophageal smooth muscle.57 The current hypothesis is that peristaltic direction and velocity result from a neural gradient along the esophagus, wherein excitatory ganglionic neurons dominate proximally and inhibitory ganglionic neurons dominate distally. This organization is supported by the demonstration with pressure topography plotting of two subsegments within the smooth muscle segment, the first of which is strongly reactive to stimulation with cholinergic drugs.58,59 The primary inhibitory neurotransmitter (formerly referred to as the nonadrenergic, noncholinergic transmitter) is nitric oxide (NO) produced from l-arginine by the enzyme NO synthase in myenteric neurons.60,61 NO synthase inhibitors reduce the latency to contraction in vivo in response to swallowing.62,63 In addition to NO neurons, there is also evidence for a role of vasoactive intestinal polypeptide (VIP)–containing neurons in the initial inhibition.64,65
Sympathectomy of the esophagus has no apparent effect on peristalsis.66,67 On the other hand, bilateral vagotomy results in paralysis of the striated muscle segment. Severing only the afferent nerve supply to the striated muscle abolishes secondary peristalsis while leaving primary peristalsis intact, highlighting the role of central programming in the latter and the necessity of afferent sensory signals in the former.2 Recordings from the cervical esophageal vagal afferents show these to be highly sensitive to intraluminal distention, implicating them as the sensory basis for secondary peristalsis. In the smooth muscle segment of the esophagus, vagal cooling or vagotomy reduces the amplitude of primary peristalsis68,69 but does not affect secondary peristalsis.70
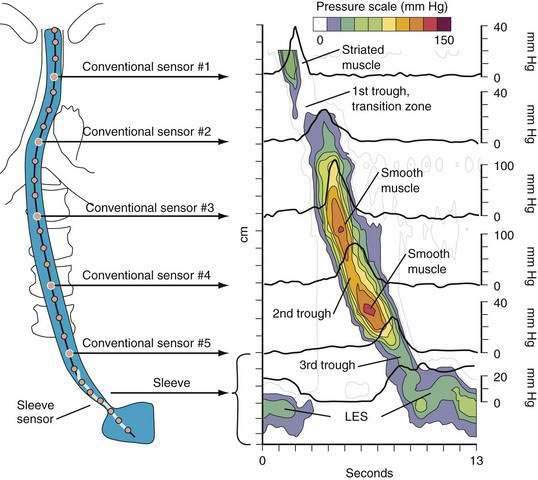
A tool introduced in the study of esophageal physiology is high-resolution manometry or, more precisely, high-resolution esophageal pressure topography. High-resolution esophageal pressure topography allows the imaging of esophageal contractile activity as a continuum not only in time but also along the length of the esophagus. Figure 42-4 illustrates the conversion from conventional manometric study to high-resolution esophageal pressure topography. Note how high-resolution esophageal pressure topography reveals that the vigor and propagation of peristalsis along the length of the esophagus is not seamless. Rather, there is a distinct transition zone between the striated and smooth muscle segments characterized by the minimal peristaltic amplitude, a slight delay in progression, and an increased likelihood of failed transmission.71 Detailed modeling studies suggest this to be the transition point between two distinct contraction waves governing the proximal and distal esophagus respectively.72 The topographic analysis also reveals a segmental characteristic of peristaltic progression through the smooth muscle esophagus with two distinct contractile segments separated by a pressure trough followed by the LES, which contracts with vigor and persistence quite dissimilar to the adjacent smooth muscle esophagus.73
The longitudinal muscle of the esophagus also contracts during peristalsis with the net effect of transiently shortening the esophagus by 2 to 2.5 cm.74 Similar to the pattern of circular muscle contraction, longitudinal muscle contraction is propagated distally as an active segment at a rate of 2 to 4 cm per second.75 The segment of contracting longitudinal muscle precedes, but overlaps with the contracting segment of circular muscle. Thus, within a given esophageal segment, the contraction of the longitudinal and circular muscle are slightly out of phase with each other. Propulsive force occurs in the zone of overlap as the delayed circular muscle contraction “catches up” with the distal longitudinal muscle contraction.75
Central mechanisms also control the contractions of esophageal longitudinal muscle. Swallowing induces peristaltic sequences with gradual activation of longitudinal muscle progressing from orad to caudad. This progression is associated with a progressive increase in latency similar to that seen with the circular smooth muscle esophagus.76 However, unlike the responses observed in the circular muscle, stimulation of decentralized vagal efferent fibers causes simultaneous contractions in the longitudinal muscle layer, suggesting this muscle layer is free of inhibitory neuron control.57
ESOPHAGOGASTRIC JUNCTION
The anatomy of the esophagogastric junction (EGJ) is complex (see Chapter 41). The distal end of the esophagus is anchored to the diaphragm by the phrenoesophageal membrane that inserts circumferentially into the esophageal musculature close to the squamocolumnar junction (SCJ). The esophagus then traverses the diaphragmatic hiatus and joins the stomach in almost a tangential fashion. Thus, there are three significant contributors to the EGJ high-pressure zone: the LES, the crural diaphragm, and the muscular architecture of the gastric cardia that constitutes the distal aspect of the EGJ high-pressure zone.
The LES is a 3- to 4-cm segment of tonically contracted smooth muscle at the distal extreme of the tubular esophagus. Surrounding the LES at the level of the SCJ is the crural diaphragm, most commonly bundles of the right diaphragmatic crus forming a teardrop-shaped canal about 2 cm long on its major axis (Fig. 42-5).77,78 Elegant anatomic studies suggest that the component of the EGJ high-pressure zone distal to the SCJ is largely attributable to the opposing sling and clasp fibers of the middle layer of gastric cardia musculature.77,79,80 In this region, the lateral wall of the esophagus meets the medial aspect of the dome of the stomach at an acute angle, defined as the angle of His. Viewed intraluminally, this region extends within the gastric lumen, appearing as a fold that has been conceptually referred to as a flap valve because increased intragastric pressure forces it closed, sealing off the entry to the esophagus.81,82
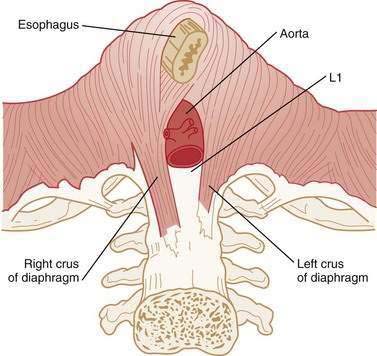
Physiologically, the EGJ high-pressure zone is attributable to a composite of both the LES and the surrounding crural diaphragm. Concurrent fluoroscopy and manometry, aided by the placement of an endoclip at the SCJ, localized the EGJ high-pressure zone as extending 1 to 1.5 cm proximal to the SCJ and about 2 cm distal to it.83 Manometric and intraluminal ultrasound studies suggest that axial asymmetry of the pressure profile is attributable to the varying thickness of the muscularis propria, whereas the radial pressure asymmetry results from asymmetrical compression by the surrounding crural diaphragm.83,84
Resting LES tone ranges from 10 to 30 mm Hg relative to intragastric pressure with considerable temporal fluctuation. The mechanism of LES tonic contraction is likely a property of the muscle itself and of nerves affecting the sphincter. This conclusion is supported by the observation that pressure within the sphincter is minimally affected by the elimination of neural activity by close intra-arterial injection of tetrodotoxin.85 Myogenic LES tone varies directly with membrane potential86 and superimposed electrical spike activity that leads to an influx of Ca++. Resting membrane potential of the LES is slightly greater (i.e., less negative) than that of the adjacent esophagus.87 Modulation of spike activity and membrane potential are not completely understood; however, it is believed that spike activity may be regulated by K+– and Ca++-activated Cl− channels.88 Sphincter tone may be maintained by inositol phosphate-mediated continuous release of intracellular calcium.89,90 Inositol phosphate concentrations are higher in the LES than in adjacent esophagus.
Apart from myogenic factors, LES pressure is also modulated by intra-abdominal pressure, gastric distention, peptides, hormones, foods, and many medications. Large fluctuations of LES pressure occur with the migrating motor complex (MMC); during phase III, LES pressure may exceed 80 mm Hg. Lesser fluctuations occur throughout the day with pressure decreasing in the post-prandial state and increasing during sleep.91 As mentioned, superimposed on the myogenic LES contraction, input from vagal, adrenergic, hormonal, and mechanical influences will alter LES pressure. Vagal influence is similar to that of the esophageal body with vagal stimulation activating excitatory and inhibitory myenteric neurons.49 Dogs provide an interesting model for studying this because they have an entirely striated muscle esophagus except for a smooth muscle band at the LES. Vagal fibers innervating the dog LES are of two types: (1) spontaneously active fibers that exhibit a sudden increase with swallowing, abruptly cease firing when the peristaltic contraction arrives, and then resume a spontaneous rate; and (2) spontaneously active fibers that cease activity with swallowing and resume normal activity when the bolus reaches the stomach.92 Thus, the LES pressure at any instant reflects the balance between excitatory and inhibitory neural input and altering the pattern of vagal discharge can result in a swallow-mediated LES relaxation. Data on humans suggest that, similar to the dog, basal LES tone is partially generated by cholinergic input.93 The excitatory and inhibitory intramural neurons are acetylcholine sensitive with nicotinic and muscarinic receptors.94 At the LES, the excitatory neurons release acetylcholine, whereas the inhibitory neurons mainly use NO as a neurotransmitter.
Adrenergic influences on LES pressure are complex and mostly mediated through actions on the myenteric neurons.34,95 Sympathetic fibers from the stellate and proximal thoracic ganglia follow the splanchnic nerve, form a recurrent loop through the celiac ganglion, and then synapse on both the excitatory and inhibitory myenteric neurons expressing α-adrenergic receptors. Adrenergic stimulation increases LES pressure by activating excitatory neurons and reducing inhibitory neuron activity. Adrenergic action on the esophageal body is the opposite of that on the LES, with direct inhibition of the muscle and inhibition of the excitatory myenteric neurons.
The crural diaphragm is also a major contributor to EGJ pressure (see Fig. 42-5). Even after esophagogastrectomy, with removal of the smooth muscle LES, a persistent EGJ pressure of about 6 mm Hg can be demonstrated during expiration.96 During inspiration there is substantial augmentation of EGJ pressure attributable to crural diaphragm contraction. Experimentally, the inspiratory augmentation of EGJ pressure can be increased even more with enhanced respiratory effort or, conversely, can be eliminated by manual ventilation. The augmentation of LES pressure observed during sustained inspiration corresponds both temporally and quantitatively with the augmentation of crural electromyographic activity, and this augmented EGJ pressure can obscure intrinsic LES relaxation induced by esophageal distention.97 Crural diaphragm contraction is also augmented during abdominal compression, straining, or coughing.98 On the other hand, during esophageal distention, vomiting, and belching, electrical activity in the crural diaphragm is selectively inhibited despite continued respiration demonstrating a control mechanism independent of the costal diaphragm.99,100 This reflex inhibition of crural activity is eliminated with vagotomy.
Lower Esophageal Sphincter Relaxation
The neural mediation of LES relaxation has been studied extensively.94,101,102 LES relaxation can be triggered by distention from either side of the esophagogastric junction or by swallowing.103 Relaxation induced by esophageal distention is modulated by bolus volume and is unaffected by vagotomy, demonstrating it to be an intramural process. Relaxation is, however, antagonized by tetrodotoxin proving that it is mediated by postganglionic nerves.104 Deglutitive LES relaxation is mediated by the vagus nerve, which synapses with myenteric plexus inhibitory neurons. Ganglionic transmission is through nicotinic and muscarinic acetylcholine receptors and can be blocked by a combination of hexamethonium (i.e., nicotinic blocker) and atropine (i.e., muscarinic blocker).
Current evidence implicates NO as the main neurotransmitter in the postganglionic neurons responsible for LES relaxation. NO is produced by NO synthase from the precursor amino acid l-arginine. Neuronal NO synthase is a soluble cytosolic enzyme and has been identified in neurons of the myenteric plexus, co-localizing with VIP, which may be a second inhibitory neurotransmitter in the LES as well as in the esophageal body.105,106 NO is released with neural stimulation in the esophagus, LES, and stomach.60,107–109 In the LES, NO has a marked inhibitory effect, and multiple in vitro and in vivo studies have shown that NO synthase inhibitors block neurally mediated LES relaxation.
Although the evidence implicating NO as the main inhibitory transmitter facilitating LES relaxation is very convincing, NO may not work alone. VIP-containing neurons have been demonstrated in the submucosal plexus and VIP relaxes the LES by direct muscle action.110–114 Electrical stimulation of LES muscle strips also causes LES relaxation and release of VIP into the muscle bath. Furthermore, VIP antiserum partially reduces LES relaxation evoked by vagal or field stimulation.65,114 It is thought that VIP acts on NO synthase containing neural terminals as a prejunctional neurotransmitter, facilitating the release of NO and on gastric muscle cells to stimulate production of NO by the muscle.115–119 In addition to VIP, there is some evidence that peptide histidine isoleucine (PHI) in the cat and, to a lesser extent, calcitonin gene-related peptide (CGRP) in the opossum may participate as inhibitory neurotransmitters.114,120 Like VIP, PHI and CGRP relax the LES by a direct action on the muscle.114,120,121 PHI is of interest because it is derived from the same precursor as VIP and coexists with VIP in the same neurons.122
The notion that multiple neurotransmitters may interact to produce LES relaxation may resolve inconsistencies and discrepancies that follow from the assumption that any one neurotransmitter is uniquely responsible for LES relaxation. Reports of the co-localization of NO synthase, VIP, pituitary adenylate cyclase activating peptide (PACAP), CGRP, and galanin in myenteric neurons of the distal esophagus support this concept.123–125
Transient Lower Esophageal Sphincter Relaxation
During rest the EGJ must contain gastric juice, but also be able to transiently relax and permit gas venting of the stomach without allowing reflux of gastric juice and food. These functions are accomplished by prolonged LES relaxations that occur transiently without swallowing or peristalsis. These transient LES relaxations (tLESRs) are thought to be an important mechanism in the pathogenesis of GERD (see Chapter 43). tLESRs are a complex reflex distinguishable from swallow-induced relaxation in several ways: (1) a prolonged (more than 10 seconds) LES relaxation independent of a pharyngeal swallowing, (2) contraction of the distal esophageal longitudinal muscle causing esophageal shortening, (3) absence of synchronized peristalsis, and (4) crural diaphragm inhibition, which is not the case with swallow-induced relaxation (Fig. 42-6).126,127 tLESRs occur most frequently in the postprandial state during gastric accommodation attributable to vagally mediated receptive relaxation of the fundus (see Chapter 48). In the setting of the completely relaxed EGJ during tLESRs, even the minimal gastroesophageal pressure gradients observed with gastric distention (3 to 4 mm Hg) are sufficient to facilitate gas venting of the stomach (belching). Thus, tLESRs are the physiologic mechanism of belching.128,129
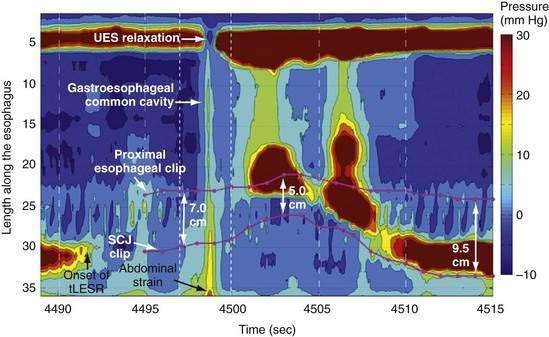
Proximal gastric distention is the major stimulus for tLESR. Distention stimulates mechanoreceptors (intraganglionic lamellar endings) in the proximal stomach activating vagal afferent fibers projecting to the nucleus of the solitary tract.127,130,131 The afferent and efferent neural pathways responsible for swallow and non-swallow LES relaxations have been compared in the mouse. The afferent arm of swallow-induced relaxation lies in the pharyngeal and superior laryngeal nerves, with the central neural circuit in the medullary subnuclei.132–135 Non–swallow-induced relaxations, in contrast, are initiated through gastric afferents in the subdiaphragmatic vagus and activate neurons in the caudal part of the dorsal motor nucleus.136 The efferent limb of both swallow and non-swallow LES relaxations lies in the preganglionic vagal inhibitory pathway to the LES. Both types of relaxation can be blocked by bilateral cervical vagotomy, cervical vagal cooling, or NO synthase inhibitors.127,137 Vagal outflow from the dorsal motor nucleus completely inhibits both the LES and the crural diaphragm, an important distinction from swallow-induced LES relaxation, which is not associated with concomitant inhibition of the crural diaphragm.
tLESRs triggered by gastric distention likely use NO and cholecystokinin (CCK) as neurotransmitters, as evidenced by increased tLESR frequency after intravenous CCK and blockade by either NO synthase inhibitors or CCK-A receptor antagonists.138–141 The CCK (and fatty meal) augmentation of tLESR frequency is mediated through CCK-A receptors.142,143 Muscarinic receptor involvement in the tLESR pathway is suggested by inhibition of tLESRs by atropine.144–147 Finally γ-aminobutyric acid (GABA)B agonists, such as baclofen, inhibit tLESRs (see Chapter 43), possibly by acting on peripheral receptors and receptors located in the dorsal motor nucleus of the vagus.148–153
ESOPHAGEAL SENSATION
The human esophagus can sense mechanical, electrical, chemical, and thermal stimuli. These stimuli are perceived as chest pressure, warmth, or pain with substantial overlap in perception among stimuli.154,155 Esophageal sensation is carried via both the vagal and spinal afferent nerves. The associated vagal neurons are located in the nodose and jugular ganglia while the corresponding spinal neurons are located in thoracic and cervical dorsal root ganglia. Vagal afferents to the upper one third of the esophagus are carried in the superior laryngeal nerve, whereas those to the remainder of the esophagus and LES are carried in vagal branches.156 The spinal afferents are contained in the thoracic splanchnic nerves projecting onto the lower cervical to upper lumbar spinal segments.156,157 Compared with vagal afferent fibers, relatively little is known about esophageal spinal afferents, but spinal pathways are thought to be primarily nociceptive. Supportive of that concept, prolonged acid perfusion produces esophageal hypersensitivity to distention via spinal sensitization.158,159 Esophageal sensations are usually perceived substernally; in the instance of pain, radiation to the midline of the back, shoulders, and jaw is very analogous to cardiac pain. These similarities are likely due to convergence of sensory afferent fibers from the heart and esophagus in the same spinal pathways, even to the same dorsal horn neurons in some cases.160
Vagal sensory endings in the esophagus consist of free nerve endings, intraganglionic laminar endings (IGLEs) within the myenteric ganglia, and intramuscular arrays (IMAs) within the muscularis propria. Labeling studies demonstrated the densest innervation of free endings between the muscularis mucosa and muscularis propria along the entire length of the esophagus.161 Electron and confocal microscopy reveal that most vagal afferents terminating in the myenteric ganglia do so in specialized laminar structure that encapsulates myenteric ganglia (IGLEs).162 One vagal afferent axon may end in several IGLEs. Combined electrophysiologic and tracer studies have demonstrated that the majority of the tension-sensitive esophageal afferents emanate from IGLEs.163 These endings detect passive and active tension of hollow viscera. In addition to IGLEs, another specialized vagal axonal ending primarily found in the longitudinal and circular smooth muscle forms a branching array parallel to the muscle fibers (IMAs).164 Although more ubiquitous in other parts of the gut, these are essentially restricted to the LES in the esophagus.165 IMAs maintain a close network with interstitial cells of Cajal and it appears that ICCs serve a trophic function.166 Functionally, IMAs appear to be stretch-sensitive endings, sensitive to changes in the muscle length.167
With IGLE and IMA sensory endings concentrated deeply within the muscularis propria beneath a relatively impermeable mucosa, it seems unlikely that intraluminal acid can directly stimulate them. However, these afferents easily respond to chemical mediators such as 5-hydroxytryptamine or alpha, beta-methylene adenosine triphosphate (ATP) as well as to mucosally applied bile or capsaicin,168,169 suggesting that these chemicals induce the release of some endogenous substance that in turn excites the muscle afferents. Supportive of that concept, muscle afferents have been shown to be sensitive to the selective purinergic P2X3 agonist alpha, beta-methylene ATP163,170 and immunohistochemical studies have documented the presence of P2X3 receptors in IGLEs implicating direct activation of purinergic P2X2 and P2X3 receptors as an initiating sensory event.171,172
With respect to free nerve endings, acid can excite esophageal vagal and spinal afferents by activating two proton-gated channels: transient receptor potential vanilloid-1 (TRPV1) and acid-sensing ion channels (ASICs).173–176 Capsaicin, a derivative of chili pepper, excites afferent fibers by activating the TRPV1 channels, which can also initiate a positive feedback loop of increased (neurogenic) inflammation via the release of neuropeptides and inflammatory substances.177,178 ASICs are the other major receptor class that are sensitive to acid, although it is doubtful that acid is their natural ligand; ASICs are probably more involved in mechanotransduction.176
Owing to its significance in the pathogenesis of reflux disease, there has been substantial interest in modulating the tLESR reflex (see Chapter 43). The current concept is that vagal afferent endings terminating in IGLEs located in the proximal stomach are primarily responsible for initiating the reflex, which is then mediated through the medulla and back to the esophagus and diaphragm via vagal efferent and the phrenic nerves.179 Pharmacologic and physiologic studies have demonstrated that the mechanotransduction properties of tension-sensitive vagal afferent fibers can be attenuated by the GABAB receptor agonist baclofen, thereby reducing the frequency of tLESR.180 Glutamate receptors are also present in vagal and spinal sensory afferent fibers, and metabotropic glutamate receptor antagonists (especially mGluR5 antagonists) have been shown to inhibit tLESR.153
ESOPHAGEAL MOTILITY DISORDERS
A working, albeit restrictive, definition of an esophageal motility disorder is an esophageal disease attributable to neuromuscular dysfunction that causes symptoms referable to the esophagus, most commonly dysphagia, chest pain, or heartburn. Employing this definition, there are relatively few firmly established primary esophageal motility disorders: achalasia, distal esophageal spasm (DES), and gastroesophageal reflux disease (GERD). GERD is clearly the most prevalent among the group and, fittingly, it is addressed in detail elsewhere in this text (see Chapter 43). Esophageal motility disorders also can be secondary phenomena in which case esophageal dysfunction is part of a more global disease: pseudoachalasia, Chagas disease, and scleroderma. Dysphagia attributable to pharyngeal or UES dysfunction can be included in a discussion of esophageal motor disorders, but this is usually as a manifestation of a more global neuromuscular disease process. The major focus of this chapter is on the primary esophageal motility disorders, particularly achalasia. However, mention is made of the secondary motility disorders and proximal pharyngoesophageal dysfunction when important unique features exist.
EPIDEMIOLOGY
Estimates of the prevalence of dysphagia among individuals older than 50 years of age range from 16% to 22%181,182 with most of this related to oropharyngeal dysfunction. Within health care institutions, it is estimated that up to 13% of hospitalized patients and 60% of nursing home residents183 have feeding problems, most of which are attributed to oropharyngeal dysfunction as opposed to esophageal dysfunction. Most oropharyngeal dysphagia is related to impaired neuromuscular function; the prevalence of the most common anatomic etiology, Zenker’s diverticulum (discussed in Chapter 23), is estimated to range from a meager 0.01% to 0.11% of the U.S. population, with a peak incidence in men between the seventh and ninth decades.184 The consequences of oropharyngeal dysphagia are severe: volume depletion, malnutrition, aspiration, choking, pneumonia, and death. In fact, mortality of nursing residents with dysphagia and aspiration can be as high as 45% over one year.185 As the U.S. population continues to age, oropharyngeal dysphagia will become an increasing problem associated with complex medical and ethical issues.
Achalasia is the most easily recognized and best-defined motor disorder of the esophagus. The annual incidence of achalasia is about 1/100,000 population in the United States and Europe,186,187 affecting both genders equally and usually presenting between ages 25 and 60.188 Because achalasia is a chronic condition, its prevalence greatly exceeds its incidence, with prevalence estimates in Europe ranging from 7.1/100,000 in Wales to 13.4/100,000 in Ireland.189 Reports of familial clustering of achalasia raise the possibility of genetic predisposition. Achalasia has been reported in one pair of monozygotic twins,190 in siblings,191 and in children of affected parents.192 However, a genetic determinant for achalasia is not strong.193 Emphasizing this point, a survey of 1012 first-degree relatives of 159 achalasic patients identified no affected relatives.194 Familial adrenal insufficiency with alacrima is a rare genetic achalasia syndrome. This condition is inherited as an autosomal recessive disease that manifests itself with the childhood onset of autonomic nervous system dysfunction including achalasia, alacrima, sinoatrial dysfunction, abnormal pupillary responses to light, and delayed gastric emptying.195 It is caused by mutations in AAAS, a gene which encodes a protein known as ALADIN.
No population-based studies exist on the incidence or prevalence of esophageal motility disorders other than achalasia. Thus, the only way to estimate the incidence or prevalence of spastic disorders is to examine data on populations at risk and reference the observed frequency of spastic disorders to the incidence of achalasia which, as detailed earlier, is about 1 per 100,000 population. Doing so, the prevalence of DES is similar to that of achalasia (or much lower if more restrictive diagnostic criteria are used). Populations at risk for motility disorders are patients with chest pain or dysphagia, so it is among these patients that extensive manometric data have been collected. Manometric abnormalities are prevalent among these groups,196–205 but in most cases the manometric findings are of unclear significance.197
PATHOGENESIS
Oropharyngeal Dysphagia
If the etiology of oropharyngeal dysphagia is not readily apparent after initial evaluation for anatomic disorders, evidence of functional abnormalities should be sought. Primary neurologic or muscular diseases involving the oropharynx are often associated with dysphagia. Thus, whereas esophageal dysphagia usually results from esophageal diseases, oropharyngeal dysphagia frequently results from neurologic or muscular diseases, with oropharyngeal dysfunction being just one pathologic manifestation. Although the specifics vary from disease to disease, the net effect on swallowing can be analyzed according to the mechanical description of the swallow outlined earlier. Table 42-1 summarizes the mechanical elements of the swallow along with the manifestation and consequence of dysfunction and provides representative pathologic conditions in which they are likely encountered. Neurologic examination may indicate cranial nerve dysfunction, neuromuscular disease, cerebellar dysfunction, or an underlying movement disorder. Functional abnormalities can be attributable to dysfunction of intrinsic musculature, peripheral nerves, or central nervous system control mechanisms. Of note, contrary to popular belief, the gag reflex is not predictive of pharyngeal swallowing efficiency or aspiration risk. The gag reflex is absent in 20% to 40% of normal adults.206
Table 42-1 Affected Phases, Manifestations, and Typical Disease Conditions Causing Oropharyngeal Dysphagia
| AFFECTED PHASE OF THE OROPHARYNGEAL SWALLOW | MANIFESTATION OF DYSFUNCTION | ASSOCIATED DISEASE(S) |
|---|---|---|
| Nasopharyngeal closure | Myasthenia gravis | |
| Laryngeal closure | Aspiration during bolus transit | |
| UES opening | ||
| Tongue loading and bolus propulsion | Sluggish misdirected bolus | |
| Pharyngeal clearance |
UES, upper esophageal sphincter.
Evident in Table 42-1, oropharyngeal dysphagia is frequently the result of neurologic or muscular diseases. Neurologic diseases can damage the neural structures requisite for either the afferent or efferent limbs of the oropharyngeal swallow. Virtually any neuromuscular disease can cause dysphagia. Because there is nothing unique to neurons controlling swallowing, their involvement in disease processes is usually random. Furthermore, in most instances, functions mediated by adjacent neuronal structures are concurrently involved. The following discussion focuses on neuromuscular pathologic processes most commonly encountered. These entities are also discussed in Chapter 35.
Stroke
Aspiration pneumonia has been estimated to inflict a 20% death rate in the first year after a stroke, and 10% to 15% each year thereafter.207 It is usually not the first episode of aspiration pneumonia, but the subsequent recurrences over the years that eventually causes death.208 The ultimate cause of aspiration pneumonia is dysphagia leading to aspiration that can occur by at least three mechanisms: absence or severe delay in triggering the swallow, reduced lingual control, or weakened laryngopharyngeal musculature.17 Conceptually, these mechanisms can involve motor or sensory impairments. Cortical strokes are less likely to result in severe dysphagia than brainstem strokes.209 Cortical strokes are also more likely to demonstrate neurologic recovery. Of 86 consecutive patients who sustained an acute cerebral infarct, 37 (43%) experienced dysphagia when evaluated within four days of the event. However, 86% of these patients were able to swallow normally two weeks later,209 with recovery resulting from contralateral areas taking over the lost function.210 Failure to recover swallowing function was more likely among patients incurring larger strokes or patients who have had prior infarcts.
Poliomyelitis
Most cases of poliomyelitis involve only the spinal cord. However, the fatality rate from bulbar polio far exceeds that of spinal disease, primarily a consequence of respiratory depression. Bulbar poliomyelitis is also associated with dysphagia. In one analysis of the persistent sequelae of bulbar poliomyelitis, 28 of 47 patients (60%) had recurrent or continued involvement of the pharynx 17 or more months after their acute illness.211 Speech and swallowing dysfunction result from weakness of the pharyngeal musculature.212 Neurologists have also observed an increasing number of patients with new paretic symptoms traceable to their remote polio infection 30 to 40 years earlier. The new, slowly progressive post-polio muscular atrophy may occur in muscles that were clinically unaffected by the acute illness.211 One investigation studied 13 patients with post-polio dysphagia and demonstrated palatal, pharyngeal, and laryngeal weakness.213 More than half of the patients evaluated demonstrated silent aspiration.
Amyotrophic Lateral Sclerosis
Amyotrophic lateral sclerosis (ALS) is a progressive neurologic disease characterized by degeneration of motor neurons in the brain, brainstem, and spinal cord. Specific symptoms are dependent on the locations of affected motor neurons and the relative severity of involvement. When the degenerative process involves the cranial nerve nuclei, swallowing difficulties ensue. Oropharyngeal dysfunction characteristically begins with the tongue and progresses to involve the pharyngeal and laryngeal musculature. Patients experience choking attacks, become volume depleted and/or malnourished, and incur aspiration pneumonia. The decline in swallowing function is progressive and predictable, invariably leading to gastrostomy feeding. Patients often die as a consequence of their swallowing dysfunction in conjunction with respiratory depression.214
Parkinson’s Disease
Although only 15% to 20% of patients with Parkinson’s disease complain of swallowing problems, more than 95% have demonstrable defects when studied videofluoroscopically.215 This disparity suggests that patients compensate in the early stages of the disease and complain of dysphagia only when it becomes severe. Abnormalities include repetitive lingual pumping prior to initiation of a pharyngeal swallow, piecemeal swallowing, and oral residue after the swallow. Patients may also exhibit a delayed swallow response and a weak pharyngeal contraction, resulting in vallecular and pyriform sinus residue. Recent data suggest this to be related to the combination of incomplete UES relaxation and a weakened pharyngeal contraction.215
Tumors
Medullary or vagal tumors are potentially debilitating with respect to swallowing. Astrocytomas are the most common tumor subtype affecting adults whereas medulloblastomas are the most common type encountered in children.216 The morbidity of these tumors is often substantially increased as a result of the relative inaccessibility of the medulla to surgery. Unilateral lesions of the vagus can result in hemiparesis of the soft palate and pharyngeal constrictors, as well as of the laryngeal musculature. Surgical manipulation of this region can even result in complete loss of the pharyngeal swallow response.217 The recurrent laryngeal nerves can be injured as a result of thyroid surgery, aortic aneurysms, pneumonectomy, primary mediastinal malignancies, or metastatic lesions to the mediastinum. Owing to its more extensive loop in the chest, the left recurrent laryngeal nerve is more vulnerable than the right to involvement with mediastinal node malignancy. Unilateral recurrent laryngeal nerve injury results in unilateral adductor paralysis of the vocal cords. This defect can result in aspiration during swallowing because of impaired laryngeal closure. It is rare, however, to have any primary pharyngeal dysfunction resultant from recurrent laryngeal nerve injury.218
Oculopharyngeal Dystrophy
Oculopharyngeal muscular dystrophy is a syndrome characterized by progressive dysphagia and ptosis. Historically, afflicted patients reaching age 50 typically died of starvation resulting from pharyngeal paralysis.219 The disease is now known to be a form of muscular dystrophy and is inherited as an autosomal dominant disorder with occurrences clustered in families of French-Canadian descent. Genetic studies of an afflicted family indicate linkage to chromosome 14, perhaps involving the region coding for cardiac alpha or beta myosin heavy chains.220 Oculopharyngeal dystrophy affects the striated pharyngeal muscles and the levator palpebrae. Although other forms of muscular dystrophy occasionally affect the pharyngeal constrictors, this is rarely a dominant manifestation. The first symptom of oculopharyngeal dystrophy is usually ptosis that slowly progresses and eventually dominates the patient’s appearance. Dysphagia may begin before, be concomitant with, or occur after ptosis. The dominant functional abnormalities are of a weak or absent pharyngeal contraction with hypopharyngeal stasis.219 Dysphagia is slowly progressive, but may ultimately lead to starvation, aspiration pneumonia, or asphyxia.
Myotonic Dystrophy
Myotonic dystrophy is a rare disorder characterized by prolonged contraction and difficulty in relaxation of affected skeletal musculature. Recent investigations suggest that even though only half of the patients complain of dysphagia, pharyngeal and esophageal motor abnormalities can be universally demonstrated. The pattern of abnormality is of a weakened pharyngeal contraction, absent peristalsis in the striated muscle esophagus, and diminished or absent peristalsis in the smooth muscle segment of the esophagus. Myotonia has not been demonstrated in any part of the esophagus.29 Thus, the risk of aspiration in this disease is similar to other forms of muscular dystrophy. Aspiration can occur during the swallow due to poor pharyngeal clearance combined with concurrent weakness of the laryngeal elevators or after the swallow when the substantial pharyngeal residue might fall into the reopened airway.
Myasthenia Gravis
Myasthenia gravis is a progressive autoimmune disease characterized by high circulating levels of acetylcholine receptor antibody and destruction of acetylcholine receptors at neuromuscular junctions. Musculature controlled by the cranial nerves is almost always involved, particularly the ocular muscles. Dysphagia is prominent in more than a third of cases and, in unusual instances, can be the initial and dominant manifestation of the disease.17 In mild cases, dysphagia may not be evident until after 15 to 20 minutes of eating. Classically, manometric studies reveal a progressive deterioration in the amplitude of pharyngeal contractions with repeated swallows. Peristaltic amplitude recovers with rest or following the administration of 10 mg edrophonium chloride, an acetylcholinesterase inhibitor. In more advanced cases, the dysphagia can be profound and associated with nasopharyngeal regurgitation and nasality of the voice, even to the extent of being confused with bulbar ALS or a brainstem stroke.221
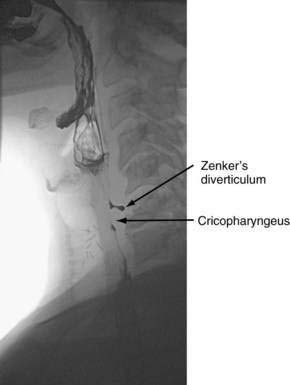
Hypopharyngeal (Zenker’s) Diverticulum and Cricopharyngeal Bar
Hypopharyngeal diverticulum and cricopharyngeal bars are closely related disease entities in that it is a cricopharyngeal bar that can result in diverticulum formation. Zenker’s diverticulum (Fig. 42-7), is discussed in Chapter 23. Zenker’s diverticulum originates in the midline posteriorly at Killian’s dehiscence, a point of pharyngeal wall weakness between the oblique fibers of the inferior pharyngeal constrictor and the transverse cricopharyngeus muscle (see Fig. 42-6).222 Other locations of acquired pharyngeal diverticula include (1) the lateral slit separating the cricopharyngeus muscle from the fibers of the proximal end of the esophagus through which the recurrent laryngeal nerve and its accompanying vessels run to supply the larynx; (2) at the penetration of the inferior thyroid artery into the hypopharynx; and (3) at the junction of the middle and inferior constrictor muscles. The unifying theme of these locations is that they are sites of potential weakness of the muscular lining of the hypopharynx through which the mucosa herniates, leading to a “false” diverticulum. The best-substantiated explanation for the development of diverticula is that they form as a result of a restrictive myopathy associated with diminished compliance of the cricopharyngeus muscle. Surgical specimens of cricopharyngeus muscle strips from 14 patients with hypopharyngeal (Zenker’s) diverticula demonstrated structural changes that would decrease UES compliance and opening.223 The cricopharyngeus samples from these patients had “fibro-adipose tissue replacement and (muscle) fiber degeneration.” Thus, although the muscle relaxes normally during a swallow, it cannot distend normally, resulting in the appearance of a cricopharyngeal indentation, or bar, during a barium swallow (Fig. 42-8). Diminished sphincter compliance necessitates increased hypopharyngeal intrabolus pressure to maintain trans-sphincteric flow through the smaller UES opening. The increased stress on the hypopharynx from the increased intrabolus pressure may ultimately result in diverticulum formation.