Chapter 7 Bile secretion and pathophysiology of biliary tract obstruction
Overview
Bile secretion is one of the major functions of the liver, and it serves two major purposes: 1) the excretion of hepatic metabolites—including bilirubin, cholesterol, drugs, and toxins—and 2) the facilitation of intestinal absorption of lipids and fat-soluble vitamins. Alterations in bile secretion may contribute to cholelithiasis (see Chapter 30) and its potential complications, such as cholecystitis (see Chapter 31) and choledocholithiasis (see Chapters 35, 36, and 37). On the other hand, obstruction of bile flow results in alterations of coagulation, the immune system, and all organ function. This chapter will discuss the physiology of bile secretion, the pathophysiology of bile obstruction, and the management of obstructive jaundice.
Bile Secretion
Bile Composition
The components of hepatic and gallbladder bile are essentially the same, but the concentration varies considerably because of the ability of the gallbladder to absorb water (Table 7.1). The gallbladder absorbs water both actively via sodium–hydrogen (Na+/H+) pumps and passively through aquaporin channels. Both chloride (Cl−) and bicarbonate (HCO3−) are absorbed by the gallbladder epithelium via the cystic fibrosis transmembrane regulator (CFTR; Swartz-Basile et al, 2007). The secretion of hydrogen ions and the absorption of bicarbonate by the gallbladder alter the acid-base balance from basic in hepatic bile to acidic in gallbladder bile.
Table 7.1 Composition of Hepatic and Gallbladder Bile
| Characteristics* | Hepatic Bile | Gallbladder Bile |
|---|---|---|
| Sodium | 160 | 270 |
| Potassium | 5 | 10 |
| Chloride | 90 | 15 |
| Bicarbonate | 45 | 10 |
| Calcium | 4 | 25 |
| Magnesium | 2 | 4 |
| Bilirubin | 1.5 | 15 |
| Protein | 150 | 200 |
| Bile acids | 50 | 150 |
| Phospholipids | 8 | 40 |
| Cholesterol | 4 | 18 |
| Total solids | — | 125 |
| pH | 7.8 | 7.2 |
Significant ranges may be seen.
* All determinations are milliequivalents per liter, except for pH.
The gallbladder mucosa also absorbs the calcium (Ca++) and magnesium (Mg++) cations. However, calcium absorption is not as efficient as the absorption of sodium and water, which leads to a significantly greater relative increase in the concentration of calcium in the gallbladder. Similarly, the concentration of bilirubin, which is not actively absorbed by the gallbladder, may be as high as 10-fold. Thus precipitation of calcium bilirubinate crystals, the major component of pigment gallstones, is much more likely to occur within the gallbladder. In addition, the biliary lipids, bile salts, phospholipids, and cholesterol all become more concentrated in the gallbladder. As the gallbladder bile becomes concentrated, several changes occur in the capacity of bile to solubilize cholesterol. The solubility in the micellar fraction is increased, but the stability of the phospholipid–cholesterol vesicles is greatly decreased. Because cholesterol crystal precipitation occurs preferentially by vesicular, rather than micellar, mechanisms, the net effect of concentrating bile is an increased tendency to form cholesterol crystals (Klein et al, 1996).
Bile Salt Secretion
Bile is secreted from the hepatocyte into canaliculi, which drain their contents into small bile ducts. Secretion of bile salts is the major osmotic force for the generation of bile flow. Bile acids are formed at a rate of 500 to 600 mg per day. The bulk of the bile salt pool is maintained in the gallbladder, followed by the liver, the small intestine, and the extrahepatic bile ducts. Bile acids are synthesized from cholesterol via two main pathways: a classic pathway leads to the formation of cholic acid, and an alternate pathway results in the synthesis of chenodeoxycholic acid. The classic pathway is the predominant mode of bile acid synthesis in humans. As a result, approximately 70% of the bile acid pool consists of cholic acid and its metabolite deoxycholic acid, with chenodeoxycholic acid occurring less commonly in human bile (Kullack-Ublick, 2004).
In plasma, bile acids circulate bound to either albumin or lipoproteins. In the space of Disse within the liver, bile salt uptake into the hepatocytes is very efficient. This process is mediated by sodium-dependent and sodium-independent mechanisms. The sodium-dependent pathway accounts for more than 80% of taurocholate uptake but less than 50% of cholate uptake (Meier & Stieger, 2002). In recent years a number of transport proteins have been identified that play a key role in this process (Fig. 7.1). The bile salt transporter is termed the sodium-taurocholate cotransporting polypeptide (NTCP), and it is exclusively expressed in the liver and located in the basolateral membrane of the hepatocyte. Sodium-independent hepatic uptake of bile acids is mediated primarily by a family of transporters termed the organic anion transporting polypeptides (OATPs). In contrast to NTCP, these transporters have a broader substrate affinity and transport a variety of organic anions, including the bile salts. OATP-C is the major sodium-independent bile salt uptake system, but OATP-A also takes up bile acids, and OATP-8 mediates taurocholate uptake.
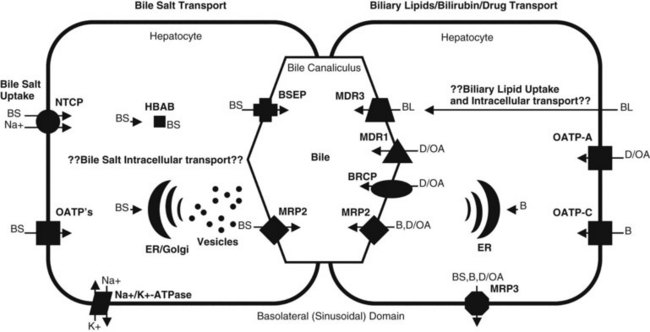
Intracellular bile acid transport occurs within a matter of seconds. Two mechanisms may be responsible for bile acid transcellular movement: one involves transfer of bile acids from the basolateral membrane to the canalicular membrane via bile acid–binding proteins (Crawford, 1996), the other moves cellular bile salt through vesicular transport. In contrast, the transport of bile salts across the canalicular membrane of hepatocytes represents the rate-limiting step in the overall secretion of bile salts from the blood into bile.
Bile salt concentrations are 1000-greater within the canaliculi than in the hepatocytes. This gradient necessitates an active transport mechanism, which is an ATP-dependent process. This bile salt export pump (BSEP) is closely related to the proteins encoded by the multidrug resistance (MDR) gene family of ATP binding cassette (ABC) transporters (Kullack-Ublick, 2004). The ABC transporters mediate the transport of metabolites, peptides, fatty acids, cholesterol, and lipids in the liver, intestines, pancreas, lungs, kidneys, brain, and in macrophages. Although BSEP is the major transporter for monovalent bile salts into the canaliculus, MDR-related protein-2 (MRP2), a member of the multidrug resistance protein family, also transports sulfated and glucuronidated bile salts into the canaliculus. MRP2 also mediates the export of multiple other organic anions, including conjugated bilirubin, leukotrienes, glutathione disulfide, chemotherapeutic agents, uricosurics, antibiotics, toxins, and heavy metals (Gerk & Vore, 2002).
Biliary Lipid Secretion
Compared with bile salts, the biliary lipids, phospholipids, and cholesterol play a secondary role in the formation of bile. Phospholipids and cholesterol are formed primarily from low-density lipoproteins circulating in plasma and from de novo synthesis by hepatocytes. Less is known about the secretion of biliary lipids compared with bile salt secretion; however, biliary lipid secretion is key for cholesterol disposal, intestinal absorption of dietary lipids, and cytoprotection against bile acid–induced hepatocyte and cholangiocyte injury (Arrese & Accatino, 2002).
Phospholipid secretion involves the delivery of phospholipids to the inner leaflet of the canalicular plasma membrane (Elferink & Groen, 2000). In humans the MDR3 transporter translocates phospholipids from the inner to the outer leaflet of the canalicular membrane. Humans with an MDR3 deficiency develop progressive familial intrahepatic cholestasis type 3 (Kullak-Ublick, 2004). These patients have no phosphatidylcholine in bile and therefore do not form mixed micelles with bile salts. As a result, toxic bile salts injure the biliary epithelium, resulting in neonatal cholestasis, cholestasis of pregnancy, and cirrhosis in adults.
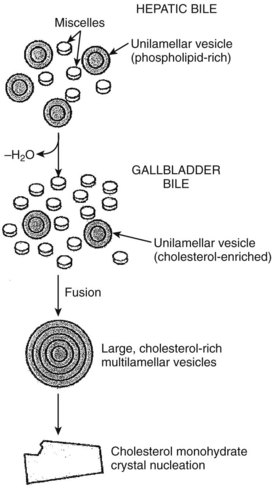
Less is known about the role of transporter proteins in cholesterol secretion, but the ABC transporters, ABCG5 and ABCG8, have been demonstrated to be involved in the elimination of plant steroids (Lee, 2004). Cholesterol is highly nonpolar and insoluble in water; thus it is insoluble in bile. The key to maintaining cholesterol in solution is the formation of micelles, a bile salt–phospholipid–cholesterol complex. Bile salts are amphipathic compounds that contain both a hydrophilic and hydrophobic portion. In aqueous solutions, bile salts are oriented with the hydrophilic portion outward. Phospholipids are incorporated into the micellar structure, allowing cholesterol to be added to the hydrophobic central portion of the micelle. In this way, cholesterol can be maintained in solution in an aqueous medium. The concept of mixed micelles as the only cholesterol carrier has been challenged by the demonstration that much of the biliary cholesterol exists in a vesicular form. Structurally, these vesicles are made up of lipid bilayers of cholesterol and phospholipids. In their simplest and smallest form, the vesicles are unilamellar, but an aggregation may take place, leading to multilamellar vesicles. Present theory suggests that in states of excess cholesterol production, these large vesicles may also exceed their capability to transport cholesterol, and cystal precipitation may occur (Fig. 7.2).
Bilirubin Secretion
In the sinusoidal membrane of the hepatocyte, bilirubin is taken up by OATP-C, a membrane transporter belonging to the OATP family (Cui et al, 2001). OATP-C is involved with the uptake of both conjugated and unconjugated bilirubin, but unconjugated bilirubin also can cross hepatic sinusoidal membranes by a diffusion process. In the hepatocyte, bilirubin binds to a driver of gluthathione-S-transferase and is catalyzed by bilirubin uridine-5-diphosphate (UDP)-glycosyltransferase to form bilirubin glucuronides. Mutations in the gene encoding bilirubin UDP-glycosyltransferase are associated with the unconjugated hyperbilirubin syndromes, Crigler-Najjar and Gilbert syndromes (Iganagi, 1998).
Enterohepatic Circulation
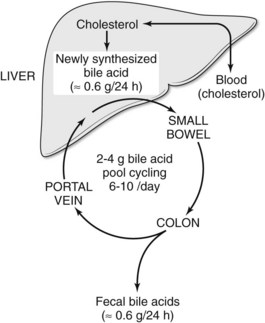
Bile salts are synthesized and conjugated in the liver; secreted into bile; stored temporarily in the gallbladder; passed from the gallbladder into the duodenum; absorbed throughout the small intestine, especially in the ileum; and returned to the liver via the portal vein. This cycling of bile acids between the liver and the intestine is referred to as the enterohepatic circulation (Fig. 7.3). The total amount of bile acids in the enterohepatic circulation is defined as the circulating bile pool. In this highly efficient system, nearly 95% of bile salts are reabsorbed. Thus, of the total bile salt pool of 2 to 4 g, which recycles through the enterohepatic cycle 6 to 10 times daily, only about 600 mg is actually excreted into the colon. Bacterial action in the colon on the two primary bile salts, cholate and chenodeoxycholate, results in the formation of the secondary bile salts, deoxycholate and lithocholate. Although some deoxycholate is reabsorbed passively by the colon, the remainder is lost in fecal waste; however, the physiology of bile salt, biliary lipid, bilirubin, bile flow, and the enterohepatic circulation are dramatically altered when the bile ducts become obstructed.
Biliary Obstruction
Causes of Jaundice
Jaundice may result from 1) increased production of bilirubin, 2) impaired hepatocyte uptake of bilirubin, 3) impaired conjugation of bilirubin, 4) impaired transport or excretion of bilirubin into the bile canaliculus, or 5) obstruction of the intrahepatic or extrahepatic biliary tree (Table 7.2
Related posts:
 Preoperative and postoperative nutrition in hepatobiliary surgery
Preoperative and postoperative nutrition in hepatobiliary surgery
 Cytokines in liver, biliary, and pancreatic disease
Cytokines in liver, biliary, and pancreatic disease
 Nonhepatic surgery in the cirrhotic patient
Nonhepatic surgery in the cirrhotic patient
 Liver transplantation for cholangiocarcinoma and other neoplastic diseases
Liver transplantation for cholangiocarcinoma and other neoplastic diseases
 Distal splenorenal shunt
Distal splenorenal shunt
 Interventional endoscopy: Technical aspects
Interventional endoscopy: Technical aspects
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


