CHAPTER 72 Liver Physiology and Energy Metabolism
LIVER CELL TYPES AND ORGANIZATION
Liver cells can be classified into three groups: parenchymal cells include hepatocytes and bile duct epithelia; sinusoidal cells are composed of hepatic sinusoidal endothelial and Kupffer cells (hepatic macrophages); and perisinusoidal cells consist of hepatic stellate cells and pit cells. Hepatocytes comprise 60% of the adult liver cell population, representing ~78% of the tissue volume (see Chapter 71).1
PARENCHYMAL CELLS
Hepatocytes
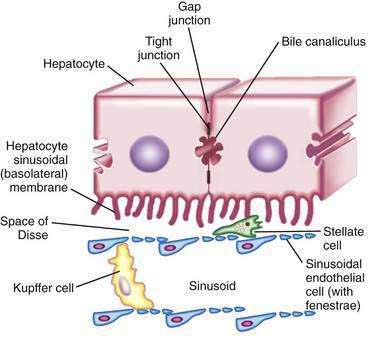
Hepatocytes are large polyhedral cells approximately 20 to 30 µm in diameter.2 Consistent with their high synthetic and metabolic activity, hepatocytes are enriched in organelles. About 30% of human hepatocytes are binucleate. Hepatocytes are polarized epithelial cells. Their plasma membranes have three distinct domains—(1) the sinusoidal surface (~37% of the cell surface) that comes in direct contact with plasma through the fenestrae of the specialized hepatic sinusoidal endothelial cells; (2) the canalicular surface (~13% of the cell surface) that encloses the bile canaliculus; and (3) contiguous surfaces. By analogy with glandular epithelia, the sinusoidal, canalicular, and contiguous plasma membrane domains are also termed basolateral, apical, and lateral surfaces, respectively.3 The sinusoidal and canalicular surfaces contain microvilli, which greatly extend the surface area of these domains.
The space between the endothelia and the sinusoidal villi is termed the space of Disse. There is a bidirectional exchange of liquids and solutes between the plasma and hepatocytes at the sinusoidal surface. In many cases, the molecular transfer is augmented by proteins that promote facilitated diffusion or energy-consuming active transport. The canalicular domains of two adjacent hepatocytes are sealed at the periphery by tight junctions (desmosomes), thereby delimiting the bile canaliculus, which is the beginning of the biliary drainage system (see Chapter 62). In contrast to the bidirectional flow at the sinusoidal surface, flow from hepatocytes into the bile canaliculi is predominantly unidirectional.
Plasma Membranes
The plasma membranes consist of lipid bilayers composed of glycerophospholipids, cholesterol, and sphingolipids that provide barrier to water and most polar substances.3,4 The inner and outer leaflets of the plasma membrane differ in lipid, protein, and carbohydrate composition, reflecting their functional differences. Protein molecules within the leaflets mediate transport of specific molecules and serve as a link with cytoskeletal structures and the extracellular matrix. Hepatocyte plasma membranes consist of 36% lipid, 54% protein, and 10% carbohydrate by dry weight. Outer leaflets of hepatocyte plasma membranes are enriched in carbohydrates.
Lipid rafts are microdomains (∼50 nm diameter) of the outer leaflets of the plasma membrane that are highly enriched in cholesterol and sphingolipids.5 These are coupled to cholesterol-rich microdomains in the inner leaflet by an unknown mechanism. Raft lipids and associated proteins diffuse together laterally on the membrane surface. Some surface receptors become associated with the rafts on ligand binding, or they can lead to “clustering” of smaller rafts into larger ones. Lipid rafts are important in signal transduction, apoptosis, cell adhesion and migration, cytoskeletal organization, and protein sorting during both exocytosis and endocytosis (see later). Certain viruses enter cells via the lipid rafts.
Membrane proteins perform receptor, enzyme, and transport functions.6 Integral membrane proteins traverse the lipid bilayer once or multiple times or are buried in the lipid. Additional “extrinsic” protein molecules are associated with plasma membrane. Membrane proteins can rotate or diffuse laterally but usually do not flip-flop from one leaflet to another. Concentration of specific membrane proteins is maintained by a balance between their synthesis and degradation by shedding of membrane vesicles, proteolytic digestion within the membrane, or internalization into the cell. Receptor proteins internalized into the cell may be degraded or recycled to the cell surface.
Cell Junctions
Hepatocytes are organized into sheets (seen as chords in two-dimensional sections) by occluding (“tight”), communicating (“gap”), and anchoring junctions (Fig. 72-1). Tight junctions or desmosomes form gasket-like seals around the bile canaliculi, thereby permitting a concentration difference of solutes between the cytoplasm and bile canaliculus. Desmosomes are specialized membrane structures that anchor intermediate filaments to the plasma membrane and link cells together. Gap junctions are subdomains of contiguous membranes of hepatocytes that comprise ~3% of the total surface membrane. They consist of hexagonal particles with hollow cores, termed connexons, made up of six connexin molecules.7 Connexons of one cell are joined to those of an adjacent cell to form a radially symmetrical cylinder that can open or close the central channel. Gap junctions are involved in nutrient exchange, synchronization of cellular activities, and conduction of electrical impulses.
Cytoskeleton
The hepatocyte cytoskeleton supports the organization of subcellular organelles, cell polarity, intracellular movement of vesicles, and molecular transport.8,9 It comprises microfilaments, microtubules, and intermediate filaments, as well as the cytoskeleton-associated proteins.10 Intermediate filaments are polymers of fibrous polypeptides (cytokeratins and lamins) that provide structural support to the cells. In addition, vimentin is expressed by hepatocytes in tissue culture, and neurofilaments appear in injured hepatocytes and form Mallory bodies (also termed Mallory-Denk bodies or Mallory’s hyaline). Hepatocytes express two cytokeratins, CK8 and CK18. Bile duct epithelial cells express these proteins and CK19. Plectin is a giant protein that cross-links intermediate filaments to each other and to the plasma membrane, microtubules, and actin filaments.
Microtubules are hollow tubular structures (with an outer diameter of 24 nm) that consist of polymerized dimers of α and β tubulin, that are involved in intracellular transport and cellular organization.11,12 Microtubules serve as tracks to the movement of cytoplasmic vesicles, mediated by adenosine triphosphatase (ATPase)-powered motor proteins, kinesin, dynein, and dynamin. Depolymerization of the microtubules, for example, by colchicine treatment, inhibits plasma protein secretion without affecting protein synthesis. Microtubules participate in cellular organization by interacting with the Golgi apparatus, intermediate filaments, and F-actin.13 They also maintain the integrity of the surface membrane during canalicular contraction.14
Microfilaments are composed of double-helical F-actin strands, which are polymers of G-actin. A large number of actin-associated proteins control the polymerization, depolymerization, and splicing of F-actin. Together with myosins, actins maintain the integrity of the cell matrix, facilitate bile canalicular contraction, and control tight junction permeability. Microfilaments are also important in receptor-mediated endocytosis and various transport processes. Collapse of the cellular structure of hepatocytes during apoptosis and formation of apoptotic bodies may be related to remodeling of the actin cytoskeleton of hepatocytes.15
Nucleus
Nuclei of hepatocytes are relatively large and have prominent nucleoli. The two concentric nuclear membranes are stabilized by networks of intermediate filaments, one inside the inner membrane and one outside the outer membrane.16 The outer nuclear membrane is in direct continuity with the ER membranes. The perinuclear space between the two nuclear membranes surrounds the nucleus and is continuous with the endoplasmic reticulum (ER) lumen. The nuclear membrane contains pores through which molecules are selectively transported to and from the cytoplasm. The ribonuclear protein (RNP) network and the perinucleolar chromatin radiate from the nucleolus.
Transport between the Nucleus and the Cytoplasm
Pores of the nuclear envelope are associated with a large number of proteins, organized in an octagonal symmetry.17 The nuclear pore complex (NPC) is a large macromolecular assembly that protrudes into both the cytoplasm and the nucleoplasm. Bidirectional nucleocytoplasmic transport occurs through the central aqueous channel in NPCs.18 Histones, DNA- and RNA-polymerases, transcription factors, and RNA-processing proteins are selectively transported into the nucleus from the cytoplasm, where they are synthesized, whereas tRNAs and mRNAs are synthesized in the nucleus and exported to the cytoplasm through the NPCs.
Endoplasmic Reticulum
The ER is the largest intracellular membrane compartment, consisting of membranous tubules or flattened sacs (cysternae) that enclose a continuous lumen or space and extend throughout the cytoplasm.19 The domain of ER in which active protein synthesis occurs has attached ribosomes and is termed the rough ER. The other domain, termed smooth ER, is devoid of ribosomes and is the site of lipid biosynthesis, detoxification, and calcium regulation. The nuclear envelope is a specialized domain of the ER.20
Golgi Complex
The Golgi complex consists of a stack of flat sac-like membranes (cysternae) that are dilated at the margins.21 Many proteins synthesized in the rough ER are transported to the Golgi apparatus in protein-filled transition vesicles. The aspect of the Golgi complex facing the ER is the cis face; the opposite side is termed the trans face. Glycoproteins are thought to be transported between the Golgi sacs via shuttle vesicles. The highly mannosylated glycosyl moiety of proteins that are N-glycosylated in the ER are processed in the Golgi sacs into mature forms. Some other proteins are O-glycosylated in the Golgi complex. These proteins are then sorted for transport to appropriate cellular organelles (see later discussion of exocytosis and endocytosis).22
Lysosomes
Lysosomes consist of a system of membrane-bound sacs and tubules that contain hydrolytic enzymes that are active at pH 4.5 to 5.23,24 The ATPase-powered proton pump maintains the acid pH by importing hydrogen ions into the lysosomal lumen.23 Lysosomal enzymes are glycoproteins with N-linked oligosaccharides. Following synthesis in the ER, the carbohydrate moieties are modified in the Golgi apparatus, where their mannose residues are phosphorylated. Recognition of these mannose 6-phosphate (M6P) groups by the M6P receptor in trans-Golgi stacks25 results in their segregation and translocation into late endosomes, which transform into lysosomes.26,27
Mitochondria
Mitochondria constitute about 20% of the cytoplasmic volume of hepatocytes and are responsible for cellular respiration.28–30 They contain the enzymes of the tricarboxylic acid cycle, fatty acid oxidation, and oxidative phosphorylation.30,31 Mitochondria conserve the energy generated by oxidation of substrates as high-energy phosphate bonds of ATP. In addition, parts of the urea cycle, gluconeogenesis, fatty acid synthesis, regulation of intracellular calcium concentration, and heme synthesis take place in the mitochondria. Mitochondria play a key role in programmed cell death, or apoptosis (see later).32
Glycolysis and fatty acid oxidation in the mitochondria generate chemical intermediates that feed into the citric acid cycle of energy-yielding reactions.33,34 The citric acid cycle breaks down acetyl coenzyme A (acetyl CoA) into three molecules of nicotinamide adenine dinucleotide (NADH), one molecule of flavin adenine dinucleotide (FADH2), and two molecules of carbon dioxide. Electrons derived from NADH and FADH2 drive an electron transport pathway in the inner mitochondrial membrane, leading to ATP production. Passage of electrons across the inner mitochondrial membrane to the space between the inner and outer membrane generates a proton gradient that drives ATP synthesis.35
Peroxisomes
Peroxisomes are spherical-appearing structures that enclose a matrix that contains a lattice or crystalline core.36 Peroxisomes are abundant in hepatocytes and are thought to be essential for life. Several oxidative catabolic reactions, as well as anabolic reactions, take place in peroxisomes, which provide important links between the metabolism of carbohydrates, lipids, proteins, fats, and nucleic acids.
Exocytosis and Endocytosis
Exocytosis and endocytosis are pathways involved in exporting, importing, and intracellular trafficking of molecules. Addition of new proteins and lipids to the plasma membrane by exocytosis and removal of membrane components into cytoplasmic compartments by endocytosis keep the cell surface in a state of dynamic polarization. During exocytosis, secreted proteins, synthesized in the ER, pass sequentially through the cis-, medial-, and trans-Golgi stacks and the trans-Golgi network and finally appear at the cell surface.37,38 This vectorial transport through the Golgi stacks occurs via vesicles that are coated by proteins termed coatamers or COP (COPI and COPII), which are distinct from clathrin (see later).39,40 Guanosine triphosphate-guanosine diphosphate (GTP-GDP) exchange factors and GTP-activating proteins that are specific for each type of vesicle stimulate membrane binding and catalytic activation of small guanosine triphosphatases (GTPases).
Once bound to the membrane, GTPases induce recruitment of COP proteins. In the ER, the first coat protein to be recruited is COPII, and vesicular/tubular clusters are formed. These clusters are thought to coalesce to form a complex tubular network, termed the ER/Golgi intermediate compartment. Acquisition of COPI proteins by the membranes of this tubular network results in the formation of vesicles that carry out bidirectional protein transport to and from the Golgi stacks. Some vesicles that emerge from the exit side of the Golgi apparatus, termed the trans-Golgi network (TGN), can transport multiple protein molecules simultaneously and release them together into the extracellular medium. Other types of vesicles that carry membrane proteins and enzymes destined for specific intracellular organelles also pass through this secretory pathway. These vesicles are sorted at the TGN, and vesicles carrying specific cargo are delivered to appropriate target organelles.41
Endocytosis is the import of extracellular macromolecules by processes that include pinocytosis, phagocytosis, receptor-mediated endocytosis (RME), and caveolar internalization.42 Pinocytosis refers to nonselective bulk-phase uptake of extracellular fluid via engulfment by plasma membrane invaginations. Phagocytosis is the ingestion of particles as well as regions of the cell surface. In contrast to these nonspecific modes of uptake, RME is a mechanism of uptake of specific molecules (ligands). After the ligands bind to their specific cell surface receptors, the ligand-receptor complexes concentrate in “pits” that are coated on the cytoplasmic surface by three-pronged structures (triskelions) composed of three heavy chains and three light chains of clathrin. The assembled coats consist of a geometric array of 12 pentagons and a variable number of hexagons, depending on the size of the coat. The coated pits pinch off into the underlying cytoplasm as coated vesicles.43 In the next step, the vesicles lose their clathrin coat and are termed endosomes. Endosomal vesicles travel along microtubules and can take three distinct pathways. Some endosomes return to the cell surface, and the contained ligand-receptor complexes are secreted out of the cells by a process termed diacytosis. Transferrin is a prototype ligand for diacytosis. Some other ligands, such as immunoglobulin A (IgA) oligomers, may traverse the cells to be secreted into bile along with the receptor. This process is termed transcytosis.44
The best studied type of RME is the classical endocytotic pathway, in which the interior of the endosome is acidified by the action of a proton pump, thereby leading to ligand-receptor uncoupling.45 By mechanisms that have not been elucidated fully, the dissociated ligands and receptors are sorted into different vesicles. The ligand-containing vesicles proceed to lysosomes, where the ligand is degraded by lysosomal hydrolases. A majority of the ligand-free receptors translocate to the cell surface and replenish the receptor pool. Some receptors, such as the insulin receptor, do not undergo recycling and are rapidly degraded in lysosomes. In addition to the recruitment of clathrin, initiation of the formation of endocytotic vesicles requires adaptor proteins, particularly AP-2, which localizes between the lipid bilayer and clathrin. Non-scaffold proteins, such as the GTPases and dynamin, are also important in the conversion of a coated pit to a coated vesicle. This function of dynamin requires association with a protein termed amphiphysin. Genetic, cell biological, and biochemical studies are identifying additional proteins that are required for clathrin coat and vesicle formation (reviewed by Stockert45). In addition to physiological ligands, many viruses use receptor-mediated endocytosis to enter cells.
Internalization via caveolae is another pathway by which macromolecules can enter cells. Binding of caveolin to the cytoplasmic aspect of cholesterol-rich lipid rafts on the plasma membrane generates 50- to 60-nm flask-shaped invaginations of the plasma membrane. These invaginations bud off into the cytoplasm to form vesicles, termed caveolae or plasmalemmal vesicles. Caveolae perform various functions, including signal transduction, calcium regulation, non-clathrin-dependent internalization, and transcytosis. Glucosyl phosphatidylinositol (GPI)-anchored proteins, the β-adrenergic receptor, and tyrosine kinase are concentrated in caveolae.46
Bile Duct Epithelial Cells
Bile duct epithelial cells, or cholangiocytes, comprise large and small subpopulations of cells, the cell volumes of which correlate roughly with the diameter of the intrahepatic bile ducts (see Chapter 62). The large cholangiocytes have a relatively more developed ER and a lower nucleus-to-cytoplasmic ratio than do the small cholangiocytes.47 The paucity of expression of cytochrome P450–dependent monooxygenase activity imparts a survival advantage to the small cholangiocytes against injury by chemicals. For example, cytochrome P450 2E1–mediated formation of toxic intermediates of carbon tetrachloride leads to the loss of large cholangiocyte function after administration of the pro-toxin, whereas the small cholangiocytes are resistant to the toxic injury.
Bile ducts are not mere passive conduits for biliary drainage but play an active role in the secretion and absorption of biliary components and regulation of the extracellular matrix composition. Cholangiocytes are highly polarized. A sodium-dependent bile salt transporter (ABAT), located at the apical (luminal) surface of cholangiocytes, mediates the uptake of conjugated bile acids by cholangiocytes, whereas an alternatively spliced truncated form of the protein (ASBT), located at the basolateral surface, mediates the efflux of the bile acids in a sodium-independent manner. The sodium-dependent glucose transporter (SGLT1), located at the apical domain, and GLUT1, a facilitative glucose transporter on the basolateral domain, are responsible for glucose reabsorption from bile. Aquaporin-1 at the apical and basolateral surfaces constitute water channels that may mediate hormone-regulated transport of water into bile by cholangiocytes. The purinergic receptor (P2u) stimulates chloride ion efflux. Activation of apical P2u by ATP, which is secreted into the bile by hepatocytes, mobilizes Ca2+ stores, thereby stimulating Cl− efflux from cholangiocytes. The large, but not the small, cholangiocytes express secretin and somatostatin receptors, the chloride/bicarbonate exchanger, and the cystic fibrosis transmembrane regulator, which may enable this population of cholangiocytes to modulate water and electrolyte secretion in response to secretin and somatostatin (see also Chapter 64).48
SINUSOIDAL CELLS
Hepatic Sinusoidal Endothelial Cells
Hepatic sinusoidal endothelial cells (HSECs) account for 20% of total liver cells. These cells are distinguished by the fenestrae (pores) in their flat, thin extensions that form sieve plates. Unlike capillary endothelial cells, HSECs do not form intracellular junctions and simply overlap each other (see Fig. 72-1B). The presence of fenestrae and the absence of a basement membrane permit plasma to enter the space of Disse and come in direct contact with the sinusoidal surface of hepatocytes.49 Diameters of the fenestrae are actively controlled by the actin-containing components of the cytoskeleton in response to changes in the chemical milieu.50 Thus, the specialized endothelial lining of hepatic sinusoids serve as a selective barrier between the blood and the hepatocytes. Hepatic endothelial cells can secrete prostaglandins and a wide variety of proteins, including interleukin (IL)-1 and IL-6, interferon, tumor necrosis factor-α (TNF-α) and endothelin.
Kupffer Cells
Kupffer cells are specialized tissue macrophages that account for 80% to 90% of the total population of fixed macrophages in the body. These cells are derived from bone marrow stem cells or monocytes and are highly active in removing particulate matter and toxic or foreign substances that appear in the portal blood from the intestine.51 Kupffer cells are located in the sinusoidal lumen and are in direct contact with endothelial cells (see Fig. 72-1). They possess bristle-coated micropinocytic vesicles, fuzzy-coated vacuoles, and worm-like structures that are special features of cells that are active in pinocytosis and phagocytosis. An abundance of lysosomes reflects their prominent role in degrading substances taken up from the bloodstream. Kupffer cells secrete a variety of vasoactive toxic mediators, which may be involved in host defense mechanisms and in pathophysiologic processes in some liver diseases. Kupffer cells increase in number and activity in chemical, infectious, or immunologic injury to the liver.52
PERISINUSOIDAL CELLS
Hepatic Stellate Cells
Hepatic stellate cells (HSCs) are also known as Ito cells, vitamin A–storing cells, fat-storing cells, or lipocytes. These cells are a part of the stellate cell system, which includes similar cells in the pancreas, lung, kidney, and intestine. Hepatic stellate cells are located between the endothelial lining and hepatocytes (see Fig. 72-1B). These mesenchymal cells represent 5% to 8% of all liver cells and are important sources of paracrine, autocrine, juxtacrine, and chemoattractant factors that maintain homeostasis in the microenvironment of the hepatic sinusoid. Microfilament and microtubule-enriched flat cytoplasmic extensions of quiescent stellate cells store vitamin A–enriched lipid droplets and spread out parallel to the endothelial lining, contacting several cells.53 HSCs express receptors for retinol-binding protein (RBP), which mediates the endocytosis of RBP-retinol complexes.54
After chronic liver injury, the slender star-shaped HSCs become activated to elongated myofibroblasts. They lose retinoids and up-regulate the synthesis of extracellular matrix components, such as collagen, proteoglycan, and adhesive glycoproteins. Stellate cell activation is the central event in hepatic fibrosis.55 Activation of HSCs is initiated by paracrine stimulation by neighboring sinusoidal endothelial cells, Kupffer cells, endothelial cells, and hepatocytes, as well as platelets and leukocytes. Endothelial cells participate in activation by producing cellular fibronectin and by converting the latent form of TGF-β to its active, profibrogenic form. Binding of TGF-β to its receptor on HSCs plays a critical role in stellate cell activation. Binding of bacterial lipopolysaccharides (LPS) arriving to the liver from the intestine to Toll-like receptor 4 (TLR4) enhances the effect of TGF-β on HSCs by two different mechanisms.56 First, increased chemokine expression by stellate cells results in chemotaxis of Kupffer cells, which secrete TGF-β. Second, LPS binding to TLR4, activates nuclear factor kappa B (NF-κB) via the adapter protein MyD88 (myeloid differentiation response protein), thereby down-regulating the TGF-β pseudoreceptor Bambi (bone morphogenic protein and the activin membrane-bound inhibitor) and thereby sensitizing the HSCs to TGF-β signaling. The three-dimensional structure of the extracellular matrix modulates the shape, proliferation, and function of HSCs, probably by signal transduction via binding to cell surface integrins, followed by changes in cytoskeleton assembly.
Pit Cells
Pit cells, the natural killer (NK) cells of the liver, are located mainly within the sinusoidal lumen, close to Kupffer cells. They have the appearance of large lymphocytes and are adherent to the sinusoidal wall, often anchored with villous extensions (pseudopods).57 In the human liver, pit cells have pronounced polarity, abundant cytoplasm containing dense granules, a conspicuous cytocenter, and a locomotory shape characterized by hyaloplasmic pseudopods and a uropod (a tail-like structure that forms on the trailing end of a moving cell). The cytoplasmic granules appear as pits by microscopy, hence the name pit cells. Pit cells are short-lived and are replenished from extrahepatic sources.
In common with circulating NK cells, the pit cells express OX-8 antigen, and some express asialoganglioside gangliotetrasylceramide (asialo-GMr1). Pit cells do not express the pan–T-cell marker, OX-19, which is expressed by circulating NK cells. Although the source of pit cells remains debated, they are antigenically related to NK cells of other viscera. Pit cells have tumor cell-killing activity in the liver and are also thought to remove virus-infected liver cells. Their per-cell cytolytic activity is greater than that of circulating NK cells. Pit cells may also have a role in controlling the growth and differentiation of liver cells and possibly in liver graft rejection.58
INTEGRATION OF THE FUNCTIONS OF THE DIFFERENT CELL TYPES
Functional integration of the various groups of liver cells occurs through direct cell-to-cell communication (e.g., via gap junctions), paracrine secretion that affects neighboring cells, cell signaling, interaction with the extracellular matrix, and generalized response to endocrine and metabolic fluxes.59 Hepatocytes and hepatic sinusoidal endothelial cells lack a continuous basement membrane, and the spatial relationship of the cells is maintained through interaction with the extracellular matrix. Anchoring to the extracellular matrix is important for the survival of hepatocytes. Anchoring also provides traction for movement and permits liver cells to receive signals from matrix components and matrix-bound growth factors. Hepatic extracellular matrix components are produced during development along the migration path of the hepatocytes and exhibit unique patterns of distribution and organization. Stellate cells, hepatocytes, and, to some extent, endothelial cells are major producers of the extracellular matrix in the liver. Excess deposition of connective tissue causes changes in hemodynamic properties and eventually impairs liver function.55
CELL-MATRIX INTERACTIONS
Cell-matrix interactions in the liver are important in maintaining hepatocyte morphology and proliferation. For example, when plated on a flat layer of collagen, hepatocytes synthesize DNA at a level that is four-fold higher than when they are grown on gels composed of basement membrane proteins. The type of matrix determines the level of expression of albumin and other hepatocyte-specific gene products in cultured hepatocytes.59,60 On the other hand, cell-cell and cell-matrix interactions determine the level of synthesis and deposition of hepatic extracellular matrix proteins by the various types of liver cells. Such interaction also modulates the production of specific enzymes and their inhibitors that mediate remodeling of the extracellular matrix.
Integrin and non-integrin receptors mediate the interaction of liver cells with extracellular matrix. Integrins bind to extracellular matrix proteins at specialized cell attachment sites that often contain the arginine-glycine-aspartate motif, thereby resulting in attachment of the extracellular matrix to the intracellular cytoskeleton network. This attachment results in changes in cell shape, spreading, and migration. Integrins also influence cell proliferation, differentiation, survival, apoptosis, and gene expression via signal transduction.61,62 Non-integrin surface receptors mediate cell attachment by different mechanisms.
COMPONENTS OF THE EXTRACELLULAR MATRIX
Components of the extracellular matrix include collagens, noncollagenous glycoproteins, and proteoglycans. The liver contains five types of collagen (I, III, IV, V, and VI) and seven classes of noncollagenous glycoproteins (fibronectin, laminin, entactin/nidogen, tenascin, thrombospondin, SPARC [secreted protein, acidic, and rich in cysteine], and undulin). Hepatic extracellular matrix also includes a large number of proteoglycans and glycosaminoglycans, such as membrane-associated syndecan, thrombomodulin, and betaglycan, and extracellular matrix-associated versican, biglycan, decorin, fibromodulin, and perlecan.59,63
REGENERATION AND APOPTOSIS OF LIVER CELLS
REGENERATION
Normal adult hepatocytes divide infrequently, with fewer than 1 in 10,000 hepatocytes undergoing mitosis at any given time—yet the liver possesses a unique capacity to replace tissue mass after liver injury or loss of liver mass. The capacity of the liver to regulate its own growth is evident in liver transplantation, where the size of the transplanted organ increases or decreases as appropriate to the size of the recipient. Such finely regulated hyperplasia of the liver is also seen after successful single-lobe liver transplantation in children.64
Hepatic regeneration has been studied extensively in rodents. Following resection of two thirds of the liver in rats, the residual liver cells proliferate and restore the liver mass within days to weeks. Although generally termed “regeneration,” this process is, in fact, restorative hyperplasia because the total liver mass, rather than the lobulated anatomic configuration, is reconstituted. In the rat, DNA synthesis peaks at 24 hours after partial hepatectomy, when approximately 35% of hepatocytes are in cell cycle. Cell division occurs six to eight hours after DNA synthesis. The time frame of DNA synthesis varies from species to species. For example, in mice, maximum DNA synthesis occurs 36 to 40 hours after hepatic resection. Because 80% to 95% of hepatocytes undergo mitosis, liver mass is restored after one or two cell divisions. All classes of hepatocytes, including diploid, tetraploid, and octaploid cells participate in this quasi-synchronized proliferation, either by mitosis of mononucleated cells or by cytokinesis of binucleated or tetranucleated hepatocytes, after DNA synthesis in all nuclei. Interestingly, adult hepatocytes, rather than liver progenitor cells, contribute to liver regeneration after partial hepatectomy. Only when the proliferation of adult hepatocytes is inhibited because of certain toxic or physical injuries do progenitor cells, often termed oval cells, proliferate. The oval cells are thought to give rise to both hepatocytes and bile duct epithelial cells.65
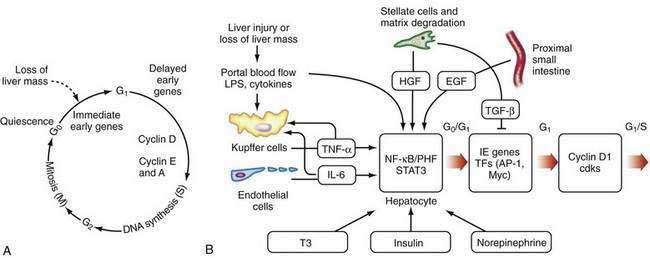
After liver injury, early signals for hepatocyte replication come from nonparenchymal cells (see Fig. 72-2B).66,67 LPS and intestine-derived cytokines stimulate Kupffer cells and hepatic sinusoidal endothelial cells to produce TNF-α and IL-6. Growth factors, such as hepatic growth factor (HGF), are released from stores in the hepatic matrix and are secreted by HSCs, whereas epidermal growth factor is secreted into portal blood by epithelial cells of the proximal small intestine and salivary glands.66 Hormones, such as triiodothyronine (T3), insulin, and nonrepinephrine, are important cooperative factors in liver regeneration.68 Replication of nonparenchymal cells lag behind that of hepatocytes by 24 to 72 hours. Initially, the newly proliferated hepatocytes form clusters, first in zone 1 and later in other zones of the liver (see Chapter 71). Regenerating endothelial cells invade these clusters and restore the single-cell thick liver plates.
Early as well as late changes occur in the expression of extracellular matrix components and the enzymes that modulate them. The mitotic phase is mostly completed in three days, and the liver mass is restituted in about seven days. Liver cells return to their quiescent state when the liver mass is restored to the original size, give or take ~10%. A balance between mitosis and apoptosis fine-tunes the restoration of hepatic mass. The strictly self-limited nature of hepatocyte replication suggests that strong regulatory pressures are present that favor replicative repression. The ability of the liver to regulate its size is dependent on signals generated outside the liver, such as hormonal or metabolic signals, as well as internal signals generated within the liver.65 Signals for cessation of growth of the regenerating liver are understood less well than those governing hepatocellular replication.
Gene Expression During Hepatic Regeneration
The regenerative process is a cascade of events that move cells from their resting G0 phase through the G1 phase, S (DNA synthesis) phase, G2 phase, and then to M (mitotic cell division) phase (Fig. 72-2A) (see Chapter 3). Expression of a large number of genes is induced or down-regulated after partial hepatectomy at transcriptional or post-transcriptional levels.66,68 The sequence of activation of various genes during liver regeneration has been elucidated by studies using partial hepatectomy and gene knockout mice that lack specific cytokines. These genes include cell cycle genes, metabolic genes, genes coding for extracellular matrix proteins, growth factors, cytokines, and transcription factors. Chronologically, these genes can be grouped into immediate early genes, delayed early genes, and cell cycle–associated genes. Expression of these genes is modulated by signal transduction pathways that receive and transduce stimuli for cell replication and tissue remodeling.
Integration of Cytokine and Growth Factors in Liver Regeneration
The early, reversible phase of liver regeneration, during which hepatocytes can enter the cell cycle by moving from the quiescent G0 state to early G1 phase, is termed priming.66 This phase is initiated by the effect of cytokines, the best studied of which include TNF-α and IL-6. Generation of reactive oxygen species as a consequence of the acute metabolic changes and release of LPS that occur in response to the loss of hepatic functional mass may have a role in triggering the initial cytokine response. During priming, NF-κB and STAT3 are activated, and AP-1 and C/EBP are expressed. Together, these factors lead to the immediate early gene expression response after partial hepatectomy (see earlier). The priming events sensitize hepatocytes to growth factors. In the absence of growth factors, the cells cannot move past a certain “restriction point” in G1.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


