CHAPTER 64 Bile Secretion and the Enterohepatic Circulation
Bile formation is essential for intestinal lipid digestion and absorption, cholesterol homeostasis, and hepatic excretion of lipid-soluble xenobiotics, drug metabolites, and heavy metals. The process of bile formation depends on hepatic synthesis and canalicular secretion of bile acids, the predominant organic anions in bile, and maintenance of hepatic bile formation is essential for normal liver function. Most of the bile acids secreted by the hepatocyte have been secreted previously into the small intestine and have undergone enterohepatic cycling. As a result, disturbances in bile acid synthesis, biliary secretion, and intestinal absorption have profound effects on hepatic and gastrointestinal physiology. Identification of the hepatic, biliary, and intestinal bile acid transporters has advanced our understanding of genetic and acquired disorders of bile formation and secretion. Transporter gene mutations have been identified in disorders such as progressive familial intrahepatic cholestasis (PFIC) types 1 to 3, intrahepatic cholestasis of pregnancy (ICP), low phospholipid-associated cholelithiasis (LPAC), Dubin-Johnson syndrome, and primary bile acid malabsorption (PBAM).1 In addition, developments in the nuclear and G-protein–coupled receptor fields have provided new insights into the regulation of bile acid synthesis, secretion, and enterohepatic cycling.2 This chapter reviews the current knowledge of the hepatic synthesis, biliary secretion, and enterohepatic circulation of bile acids.
Bile is a complex, lipid-rich micellar solution that is isosmotic with plasma and composed primarily of water, inorganic electrolytes, and organic solutes such as bile acids, phospholipids (mostly phosphatidylcholine), cholesterol, and bile pigments (Table 64-1). The relative proportions of the major organic solutes in bile is illustrated in Figure 64-1.The volume of hepatic bile secreted is estimated to range from 500 to 600 mL per day, and bile acids are the dominant organic components. Actively secreted across the canalicular membrane, bile acids induce the secretion of other biliary constituents. In healthy humans, canalicular secretion is efficient and remarkably concentrative; the intracellular monomeric concentration of bile acid is less than 10 mmol/L in the hepatocyte and more than 1000 mmol/L in canalicular bile. Bile acids travel down the biliary tree and are stored in the gallbladder. After a meal the gallbladder contracts and empties its contents into the duodenum, where bile acids facilitate absorption of cholesterol and fat-soluble vitamins. Bile acids are poorly absorbed in the proximal small intestine but are absorbed almost completely by the terminal ileum. The bile acids are returned to the liver in the portal circulation, actively absorbed at the hepatocyte sinusoidal membrane, and re-secreted into bile.3
Table 64-1 Composition of Hepatic Bile
| COMPONENT | CONCENTRATION |
|---|---|
| Electrolytes and minerals (mmol/L): | |
| Sodium | 140-160 |
| Potassium | 3-8 |
| Chloride | 70-120 |
| Bicarbonate | 20-50 |
| Calcium | 1-5 |
| Phosphate | 0-1.2 |
| Magnesium | 1-3 |
| Metals (µmol/L): | |
| Iron | 2-72 |
| Copper | 12-21 |
| Organic constituents (mmol/L): | |
| Bile acids | 5-50 |
| Bilirubin (total) | 1-2 |
| Phospholipid (lecithin) | 0.5-20.0 |
| Cholesterol | 0.5-1.0 |
| Glutathione | 3-5 |
| Glucose | 0.2-1.0 |
| Urea | 2.2-6.5 |
| Protein (g/dL) | 0.2-3.0 |
Values obtained from measurements of human bile are drawn from Albers CJ, Huizenga JR, Krom RA, et al. Composition of human hepatic bile. Ann Clin Biochem 1985; 22:129-32; Keulemans YC, Mok KS, de Wit LT, et al. Hepatic bile versus gallbladder bile: A comparison of protein and lipid concentration and composition in cholesterol gallstone patients. Hepatology 1998; 28:11-16; and Ho KJ. Biliary electrolytes and enzymes in patients with and without gallstones. Dig Dis Sci 1996; 41:2409-16.
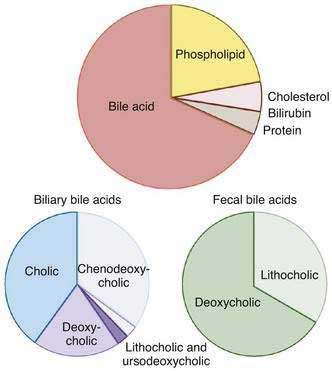
Figure 64-1. Top, Typical solute composition of hepatic and gallbladder bile in healthy humans. Bile acids are the primary solute in bile, constituting approximately 67% of bile. Phospholipid accounts for approximately 22%, cholesterol 4%, bilirubin 0.3%, and protein 4.5% of bile. Biliary bile acid composition is shown in the bottom left. Cholic acid, chenodeoxycholic acid, and deoxycholic acid constitute more than 95% of the biliary bile acids, and virtually all of the biliary bile acids are in conjugated form. The proportion of biliary lithocholic acid and ursodeoxycholic acid varies but rarely exceeds 5%. The majority of lithocholic acid in bile is present in sulfated form. When administered in therapeutic doses, the proportion of ursodeoxycholic acid in bile may rise to as much as 40%. Fecal bile acid composition is shown in the bottom right. Fecal bile acids are almost entirely unconjugated as a result of bacterial deconjugating enzymes in the colon and consist primarily of the dehydroxylated bile acids, deoxycholic acid, and lithocholic acid (see Figure 64-2).
The functions of bile acids in the liver and gastrointestinal tract are multiple.3,4 First, bile acids induce bile flow and hepatic secretion of biliary lipids (phospholipid and cholesterol). The vectorial movement of bile acids from blood into the bile canaliculus generates an osmotic water flow and is a major determinant of bile formation. Second, bile acids play an important role in the digestion of dietary fats and are essential for the intestinal absorption of cholesterol and fat-soluble vitamins. Bile acids promote intestinal absorption by solubilizing dietary lipids and lipid digestion products as mixed micelles to facilitate their aqueous diffusion across the intestinal mucosa. Fat-soluble vitamins (A, D, E, and K) are poorly absorbed in the absence of bile acid micelles, and disturbances in the secretion or enterohepatic cycling of bile acids lead to fat-soluble vitamin deficiency. Along with their major role in promoting dietary lipid absorption, bile acids may facilitate intestinal absorption of protein by accelerating protein hydrolysis by pancreatic proteases.5 Third, bile acids play a complex role in maintaining cholesterol homeostasis. On one hand, bile acids promote cholesterol intake by facilitating the intestinal absorption of biliary and dietary cholesterol. On the other hand, bile acids function through several mechanisms to promote cholesterol elimination from the body. Bile acids are water-soluble end products of cholesterol catabolism and a major route for the elimination of cholesterol via fecal excretion. Bile acids also promote hepatic secretion of cholesterol into bile by inducing bile flow and solubilizing biliary cholesterol, thereby enabling cholesterol to move from the hepatocyte to the intestinal lumen for elimination. Fourth, bile acids contribute to the antimicrobial defense mechanisms of the intestine through direct bacteriostatic actions of bile acid–fatty acid mixed micelles in the proximal small intestine and by inducing expression of antimicrobial genes in the distal small intestine.6,7 Fifth, bile acids act to prevent the formation of calcium gallstones and oxalate kidney stones.8 Sixth, bile acids can function as hormones to regulate the enterohepatic circulation as well as fat, glucose, and energy homeostasis by signaling through nuclear and G-protein–coupled receptors.2
BILE ACID SYNTHESIS AND METABOLISM
Bile acids are synthesized from cholesterol in pericentral hepatocytes of the hepatic acini. In this process, cholesterol, a lipophilic compound, is converted into a water-soluble product. In humans the newly synthesized (primary) bile acids are cholic acid (CA), a trihydroxy-bile acid with hydroxy groups at the C-3, C-7, and C-12 positions, and chenodeoxycholic acid (CDCA), a dihydroxy-bile acid with hydroxy groups at the C-3 and C-7 positions (Fig. 64-2). The kinetics for bile acid synthesis and turnover in humans are summarized in Table 64-2. Hepatic bile acid synthesis involves two major pathways, the “classical” neutral pathway (cholesterol 7α-hydroxylase pathway) that favors CA biosynthesis and the “alternative” acidic pathway (oxysterol 7α-hydroxylase pathway) that favors CDCA biosynthesis.9 In the classical pathway, the enzyme cholesterol 7α-hydroxylase (cytochrome P-450 7A1 [CYP7A1]) converts cholesterol directly into 7α-hydroxycholesterol. In the alternative pathway, cholesterol must first be converted by C-24, C-25, or C-27 sterol hydroxylases into an oxysterol, the major species being 27-hydroxycholesterol, before conversion by the oxysterol 7α-hydroxylase (CYP7B1).
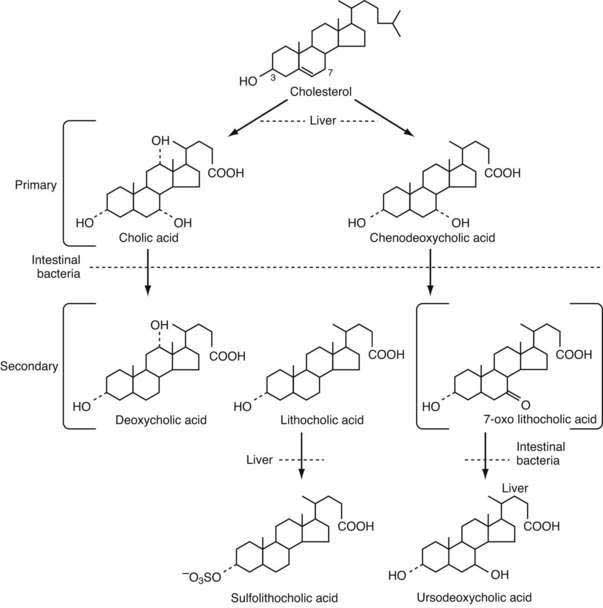
Table 64-3 Proteins Involved in the Regulation of Bile Acid Synthesis and Enterohepatic Cycling
| PROTEIN (GENE) | DESCRIPTION AND FUNCTION OF PROTEIN IN BILE ACID METABOLISM |
|---|---|
| FXR (NR1H4) | Bile acid–activated nuclear receptor; regulation of bile acid synthesis, transport, and metabolism |
| HNF4α (NR2A1) | Nuclear receptor; positive regulator of cytochrome P-450 7a1 (CYP7a1) expression and hepatic bile acid synthesis |
| SHP (NR0B2) | Nuclear receptor; negative feedback regulation of hepatic bile acid synthesis by antagonizing HNF4α, LRH-1; regulation of bile acid transport and metabolism |
| LRH-1 (NR5A2) | Nuclear receptor; positive regulator of CYP7a1 expression and hepatic bile acid synthesis |
| PXR (NR1I2) | Bile acid and xenobiotic-activated nuclear receptor involved in detoxification of secondary bile acids |
| VDR (NR1I1) | Vitamin D and bile acid–activated nuclear receptor; involved in detoxification of LCA |
| FGFR4 (FGFR4) | Membrane receptor; negative feedback regulation of CYP7a1 and hepatic bile acid synthesis |
| β-klotho (KLB) | Membrane co-receptor associated with FGFR4; confers liver specificity to FGFR4-FGF19 pathway; negative feedback regulation of CYP7a1 and hepatic bile acid synthesis |
| FGF19 (FGF19) | Protein growth factor; secreted by intestine in response to bile acids; regulates hepatic bile acid synthesis via FGFR4:β-klotho |
FXR, farnesoid X-receptor; FGF19, fibroblast growth factor 19; FGFR4, fibroblast growth factor receptor 4; HNF4α, hepatocyte nuclear factor 4 alpha; LCA, lithocolic acid; LRH-1, liver receptor homolog 1; PXR, pregnane X-receptor; SHP, small heterodimer partner; VDR, vitamin D receptor.
The overall process of bile acid biosynthesis is complex, involving 17 different enzymes divided into two broad groups.9 The first group performs modifications to the sterol ring structure, whereas the second group modifies the sterol side chain. In the classical pathway, sterol ring modifications precede side chain changes, whereas in the alternative pathway the side chain modifications occur before or during changes to the sterol ring structure. Of the two major biosynthetic pathways, the classical (CYP7A1) pathway is quantitatively more important in humans. This conclusion is supported by the finding that bile acid production is decreased by almost 90% in an adult patient with an inherited mutation in the CYP7A1 gene.10 In contrast to adults, the alternative pathway may be dominant in neonates, as evidenced by the low expression of CYP7A1 in newborns and the finding of severe cholestatic liver disease in an infant with an inherited CYP7B1 mutation.11
The rate-limiting step for the classical pathway is the enzyme CYP7A1. Bile acid feedback inhibition of CYP7A1 is well established experimentally; bile acid synthesis is decreased after administration of hydrophobic bile acids and increased by interruption of the enterohepatic circulation following ileal resection or administration of bile acid sequestrants.12 In contrast to the classical (CYP7A1) pathway, the alternative pathway for bile acid synthesis is not subject to regulation by bile acids but is controlled by cholesterol delivery to the mitochondria, the site of 27-hydroxylation.13 The molecular mechanisms responsible for the negative feedback regulation of the CYP7A1 pathway involve the liver and small intestine and have been elucidated. For the major pathway, bile acids act as ligands for the farnesoid X receptor (FXR) in ileal enterocytes to induce synthesis of an endocrine hormone, the fibroblast growth factor-19 (FGF19). FGF19 is secreted into the portal circulation and acts on hepatocytes through its cell surface receptor, a complex of the β-klotho protein and fibroblast growth factor-4, to repress CYP7A1 expression and bile acid synthesis.14–16 In a second pathway, excess bile acids in the hepatocyte activate FXR and indirectly suppress expression of CYP7A1 through a complex mechanism that involves the orphan nuclear receptor designated small heterodimer partner (SHP). SHP interferes with the activity of several nuclear receptors including hepatocyte nuclear factor 4α (HNF4α) and liver receptor homolog 1 (LRH-1), which are required for expression of the CYP7A1 gene.12,17 Finally, bile acids also inhibit CYP7A1 gene transcription in a SHP and FGFR4-independent fashion through activation of the c-Jun NH2-terminal kinase (JNK) pathway.18,19 These complex molecular titrations link bile acid synthesis to changes in intestinal as well as hepatic bile acid levels. The receptors and protein factors that participate in the regulation of bile acid synthesis and the enterohepatic circulation of bile acids are summarized in Table 64-3.
Before secretion into the bile canaliculus, both CA and CDCA are N-acyl amidated with glycine or taurine, a process commonly termed conjugation. This conjugation enhances the hydrophilicity of the bile acid and the acidic strength of the side chain, in essence converting a weak acid (pKa ≈ 5.0 for the unconjugated bile acid) to a strong acid (pKa ≈ 3.9 for the glycine conjugate; pKa ≈ 2.0 for the taurine conjugate). The major function of conjugation is to decrease the passive diffusion of bile acids across cell membranes during their transit through the enterohepatic circulation. As a result, conjugated bile acids are absorbed only if a specific membrane carrier is present. Compared with unconjugated bile acids, conjugated bile acids are also more soluble at acidic pH and more resistant to precipitation in the presence of high concentrations of calcium. The net effect of conjugation is to maintain high intraluminal concentrations of bile acids down the length of the small intestine to facilitate lipid digestion and absorption. The importance of bile acid conjugation is illustrated by the finding that inherited defects in bile acid conjugation present with the clinical phenotype of fat-soluble vitamin malabsorption and steatorrhea.11,20
Most of the conjugated bile acids secreted into the small intestine are efficiently absorbed intact; however, bile acids are also metabolized during their passage down the intestine, by the endogenous bacterial flora. In the distal small intestine, the bacterial flora deconjugates a small fraction, ≈15%, of the bile acids. The unconjugated bile acids are absorbed passively or actively and returned to the liver, where they are efficiently reconjugated and mix with newly synthesized bile acids to be re-secreted into bile. This process of intestinal deconjugation and hepatic reconjugation is a normal part of bile acid metabolism. An additional bacterial modification is the epimerization of the 3α-hydroxy or 7α-hydroxy groups to their corresponding 3β- or 7β-hydroxy forms.21,22 The 7α-hydroxy group of CDCA is epimerized to form the 3α,7β-dihydroxy bile acid, ursodeoxycholic acid (UDCA). After being absorbed from the intestine, UDCA is conjugated to taurine or glycine in the liver and circulates as a minor component, normally less than 5%, of the bile acid pool. In addition to endogenous formation, UDCA is used as a therapeutic agent in various forms of cholestatic liver disease (see Chapters 76 and 89).23
A small fraction of bile acids secreted into the small intestine escapes absorption and passes into the large intestine, where deconjugation is almost complete. In the colon, the action of bacterial 7α-dehydroxylase converts CA to deoxycholic acid (DCA), a dihydroxy bile acid with hydroxy groups at the C-3 and C-12 positions, and converts CDCA to lithocholic acid (LCA), a monohydroxy bile acid with a hydroxy group at position C-3 (see Fig. 64-2).21 The colon absorbs about 50% of the DCA formed as well as a small fraction of the LCA. After returning to the liver, DCA is reconjugated and circulates with the primary bile acids. Hepatic conjugation of the circulating bile acids is extremely efficient, so virtually all the biliary bile acids (primarily CA, CDCA, DCA, and UDCA) are in conjugated form. Bacterial deconjugation and dehydroxylation in the colon are also efficient, so the feces contain mainly the unconjugated secondary bile acids DCA and LCA (see Fig. 64-1).
The secondary bile acids can be metabolized further by various pathways, including hepatic re-epimerization of 3β-hydroxy bile acids, hepatic reduction of 7-oxo-lithocholate to CDCA or UDCA, and sulfation of bile acids by the liver or kidney (see Fig. 64-2). Additional minor pathways include hydroxylation and glucuronidation of bile acids. The conjugation of LCA with sulfate or glucuronide blocks intestinal absorption, and the modified LCA is rapidly lost from the circulating pool of bile acids. In humans, sulfation of LCA plays an important protective role, because unmodified LCA is intrinsically hepatoxic.24 The molecular mechanisms responsible for inducing these hepatoprotective modifications are being elucidated and involve induction of cytosolic sulfotransferases and cytochrome P-450 enzymes by the nuclear receptors pregnane X receptor (PXR) and constitutive androsterone receptor (CAR).25 For example, LCA, as well as other xenobiotic inducers such as rifampin, bind and directly activate PXR, thereby stimulating important mechanisms for hepatic detoxification and transport of bile acids.26
THE ENTEROHEPATIC CIRCULATION
The anatomic components of the enterohepatic circulation are the liver, biliary tract, gallbladder, small intestine, portal venous circulation, and, to a lesser extent, colon, systemic circulation, and kidneys (Fig. 64-3). At a fundamental level, the enterohepatic circulation of bile acids can be considered to consist of a series of storage chambers (gallbladder, small intestine), valves (sphincter of Oddi, ileocecal valve), mechanical pumps (bile canaliculi, biliary tract, small intestine), and chemical pumps (hepatocyte, cholangiocyte, and ileocyte).
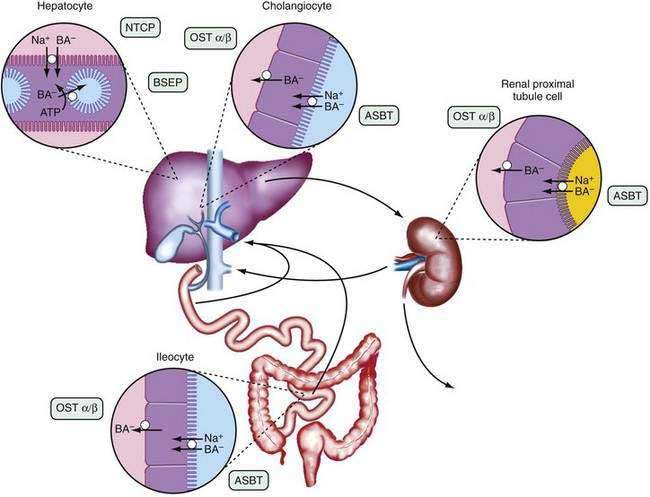
Efficient intestinal reabsorption and hepatic extraction of bile acids permit an effective recycling and conservation mechanism that largely restricts bile acids to the intestinal and hepatobiliary compartments. During fasting, bile acids move down the biliary tract and are concentrated approximately 10-fold in the gallbladder. After an overnight fast, most of the bile acids are sequestered in the gallbladder, resulting in low levels of bile acids in the small intestine, portal vein, systemic circulation, and liver. In response to a meal, cholecystokinin is released from the intestinal mucosa and acts on the biliary tree to relax the sphincter of Oddi and stimulate gallbladder contraction. A concentrated solution of mixed micelles (bile acids, phospholipids, and cholesterol) is emptied from the gallbladder via the bile duct into the small intestine, where the micelles facilitate fat absorption by stimulating the action of pancreatic lipase on triglyceride, solubilizing the hydrolytic products, and delivering lipids to the mucosal surface. During the digestion of a large meal, the gallbladder remains contracted, and bile acids secreted by the liver bypass the gallbladder and empty directly into the duodenum. During this period, the bile acid concentration in the small intestine is approximately 5 to 10 mmol/L; micelle formation requires that the intraluminal bile acid concentration be greater than 1.5 mmol/L. During the interdigestive period, the sphincter of Oddi contracts and the gallbladder relaxes, causing a larger fraction of the secreted bile acid to enter the gallbladder for storage. This gallbladder relaxation is mediated in part by the ileal release of fibroblast growth factor 19 (FGF19).27 Therefore, the enterohepatic cycling of bile acids accelerates during digestion and slows between meals and during overnight fasting. This rhythm of bile acid secretion is maintained even after cholecystectomy.28 When the gallbladder is absent, bile acids are stored in the proximal small intestine. After ingestion of a meal, small intestinal contractions propel the stored bile acids to the distal ileum, where they are actively reabsorbed.
The enterohepatic cycling of bile acids is extremely efficient; less than 10% of the intestinal bile acids escape reabsorption and are eliminated in the feces. Bile acids are absorbed from the small intestine predominantly by an active transport system restricted to the terminal ileum and, to a lesser extent, by passive absorption down the length of the intestine. Of all the conjugated anions secreted into bile, only bile acids are actively absorbed in conjugated form and undergo an enterohepatic circulation. In adult humans, the enterohepatic circulation maintains a bile acid pool size of 50 to 60 mmol per kg body weight, corresponding to approximately 2 to 4 g. The bile acid pool cycles two to three times per meal, resulting in 6 to 10 cycles per day, and the intestine may reabsorb between 10 and 30 g of bile acid per day. Approximately 0.2 to 0.6 g of bile acids escapes reabsorption and is eliminated in the stool each day. Hepatic conversion of cholesterol to bile acid balances fecal excretion, and this process represents an important route for elimination of cholesterol from the body. The kinetics of bile acid turnover in humans are summarized in Table 64-2.
HEPATIC BILE ACID TRANSPORT AND BILE SECRETION
Hepatic adenosine triphosphate (ATP)–dependent carriers actively secrete bile acids into the canalicular lumen, from which they are too large to diffuse back across the paracellular junctions that line the canaliculi. Solutes such as the conjugated bile acids that are actively pumped across the canalicular membrane generate bile flow and are termed primary solutes. The list of primary solutes includes conjugated bilirubin, glutathione, heavy metals, and conjugates of various metabolites and xenobiotics. Water, plasma electrolytes, calcium, glucose, amino acids, bicarbonate, and other low-molecular-weight solutes that flow into the canaliculus in response to the osmotic gradient are termed secondary solutes. The choleretic activity of each primary solute is defined as the amount of bile flow induced per amount of solute secreted. The choleretic activity varies for different bile acid species and ranges from 8 to 30 microliters of bile flow induced per micromole of bile acid secreted. In humans most canalicular bile flow is generated by bile acid secretion; however, the secretion of other primary solutes by the hepatocyte and biliary epithelium also contributes to bile formation. Newly secreted hepatic canalicular bile is modified during its transit in the biliary tract via the action of ductule epithelial cells (cholangiocytes), and this ductular secretion may account for up to 40% of the volume of bile. The ductular modifications to hepatic bile include the absorption of solutes such as glucose, amino acids, and bile acids; the secretion of solutes such as bicarbonate and chloride; and the movement of water through specific channels (aquaporins) and paracellularly.29,30
BILE ACID–INDEPENDENT BILE FLOW
Canalicular bile flow is generated by active secretion of primary solutes in addition to bile acids. Reduced glutathione (GSH) and bicarbonate (HCO3−) constitute the major components of the bile acid–independent fraction of bile flow. The ATP-dependent canalicular secretion of GSH and GSH conjugates via the multidrug resistance–associated protein 2 (MRP2) plays a particularly important role. In addition to being secreted at high concentrations into bile, intraluminal catabolism of GSH by gamma glutamyl transpeptidase (GGTP) further raises the solute concentration and contributes to the osmotic driving force for canalicular bile formation. Besides the ATP-dependent secretion of organic anions into bile, hepatic and biliary ATP-independent secretion of bicarbonate via the HCO3−/Cl− anion exchanger AE2 contributes to the bile acid–independent bile flow.31 The majority of this HCO3− secretion occurs at the level of the bile duct epithelial cells in response to stimulation by a variety of hormones and neuropeptides, such as secretin and vasoactive intestinal peptide.32
CHOLEHEPATIC SHUNT PATHWAY
The term cholehepatic shunt was coined to describe the cycle whereby unconjugated dihydroxy bile acids secreted into bile are absorbed passively by cholangiocytes, returned to the hepatocyte via the periductular capillary plexus, and resecreted into bile. Absorption of the protonated unconjugated bile acid molecule generates a bicarbonate anion, resulting in a bicarbonate-rich choleresis. Premature absorption and resecretion of the bile acid also increases bile acid-dependent bile flow. This cycle explains the hypercholeresis observed for unconjugated C-24 dihydroxy bile acids such as UDCA, unconjugated C-23 bile acid analogs such as norursodeoxycholate (norUDCA), and certain drugs such as the nonsteroidal anti-inflammatory drug sulindac.33






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


