Stomach: Normal Structures and Developmental Abnormalities
EMBRYOLOGY
Stomach
The stomach appears at 14 weeks as a fusiform dilation of the caudal part of the foregut. The dorsal border grows faster than the ventral border, establishing the greater curvature of the stomach. As the stomach acquires its adult shape, it rotates 90 degrees in a clockwise direction on its longitudinal axis. The adult orientation of the stomach is established as the ventral border (lesser curvature) moves to the right, the dorsal border (greater curvature) moves to the left, the original left side becomes the ventral surface, and the original right side becomes the dorsal surface. These changes explain why the left vagus nerve supplies the anterior (ventral) wall of the adult stomach and the right vagus innervates the posterior (dorsal) wall (Fig. 12-1).1
The epithelial lining and gastric glands develop from foregut endoderm. Splanchnic mesoderm produces gastric smooth muscle, the lesser omentum, and the dorsal mesentery (dorsal mesogaster and greater omentum). Gastric epithelial cells express a range of peptide hormones known to regulate gastric functions including digestive enzymes, mucus, and hormones that regulate gastric motility. At 8 weeks, the developing human stomach has gastrin-containing cells in the antrum, and somatostatin cells in both the antrum and the fundus. At 10 weeks, glucagon-containing cells are seen the gastric fundus; these are preceded by glicentin (enteroglucagon—the precursor of glucagon) but have disappeared postnatally, although occasional glicentin or glucagon-containing cells can be found in various poly and cancers.2 Serotonin-containing cells are seen in the antrum and fundus by 11 weeks.3, 4, 5, 6
Cardia
There has been considerable controversy regarding whether the cardia is normal or is acquired in response to reflux.7, 8 In embryos, there can be either a direct transition from oxyntic to esophageal squamous mucosa or a transition from oxyntic to cardiac to esophageal7, 9 suggesting that cardia is physiological and may even develop in response to gastroesophageal reflux in utero. The issue assumes potential importance in adults, where a direct oxynto-squamous transition is rare, so that an argument can be made for the cardia being acquired in at least some adults. While this should be analogous to how much gastric metaplasia is present in the first part of the duodenum, which likely also develops in response to acid reaching that part of the duodenum, it assumes potential importance in the esophagus if it is acquired in response to gastroesophageal reflux disease. This could potentially be interpreted as acquired and therefore to have potential for neoplastic transformation—with or without goblet cells. It can therefore be argued that, unless this starts acquiring intestinal mucosa features, it likely has little risk.10 However, this is a moving target, as in some studies intestinal metaplasia can be present in over a third of the population if looked for.11 We therefore suspect that normal cardia has no increased risk of neoplasia.
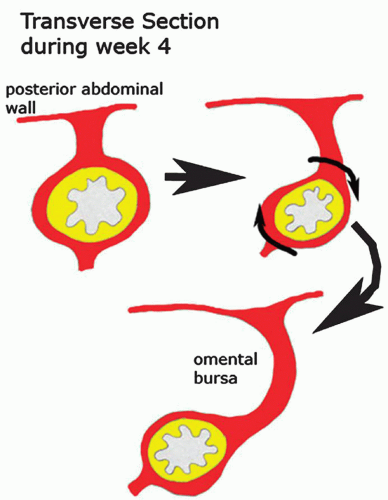 Figure 12-1. The adult orientation of the stomach is established as the ventral border (lesser curvature) moves to the right, the dorsal border (greater curvature) moves to the left, the original left side becomes the ventral surface, and the original right side becomes the dorsal surface. |
Duodenum
The duodenum develops from the most caudal part of the foregut and the most cranial part of the midgut. These parts grow rapidly and form a C-shaped loop that projects ventrally. The junction of the foregut and the midgut is at the apex of this embryonic duodenal loop usually immediately below the ampulla of Vater. Because of its derivation from both the foregut and the midgut, the duodenum is supplied by branches of the celiac and superior mesenteric arteries. During the 5th and 6th weeks, the duodenal lumen is reduced and may be obliterated by epithelial cells, but it recanalizes by the end of the embryonic period (8th week).1 Most of the duodenal ventral mesentery disappears, but the free border remains and forms the ventral border of the epiploic foramen.
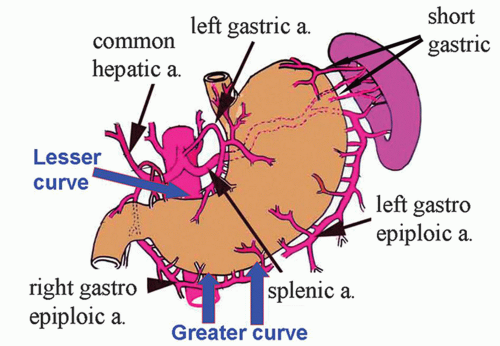 Figure 12-2. Outline of the stomach with its rich blood supply from all three branches of the celiac trunk, namely the left gastric, splenic, and common hepatic arteries (a. = artery). |
NORMAL STRUCTURE OF THE STOMACH
Anatomy
The stomach is J-shaped, although there is considerable variation, depending on the degree of distention and the body habitus. Between its two areas of fixation at each end, the stomach is quite mobile. It is fixed above at the esophagogastric junction and below at the gastroduodenal junction. The two curvatures of the stomach are designated the lesser and greater curvatures (Fig. 12-2). Externally, the stomach is covered completely by peritoneum, except where the blood vessels run along its curvatures (Fig. 12-2), and a small bare area posterior to the cardiac orifice. The peritoneum is reflected at the lesser curvature forming the lesser omentum that extends to the liver. Likewise, the peritoneum is reflected at the greater curvature to become the greater omentum (a double layer of fatty peritoneum suspended from the greater
curvature). The greater omentum is an apron-like structure hanging off the stomach that has a remarkable ability to stick to damaged or perforated parts of the gastrointestinal tract, sealing off leaks and giving some protection against peritonitis. Excess fat may be stored on the greater omentum, especially in men (hence beer belly).12
curvature). The greater omentum is an apron-like structure hanging off the stomach that has a remarkable ability to stick to damaged or perforated parts of the gastrointestinal tract, sealing off leaks and giving some protection against peritonitis. Excess fat may be stored on the greater omentum, especially in men (hence beer belly).12
Gastric surfaces (relations of the stomach). The antero-superior surface of the stomach is in contact with the diaphragm (fundal region), gastric surface of the spleen, left and quadrate lobes of the liver, the anterior abdominal wall, and the transverse colon— when the stomach is empty. The postero-inferior surface (stomach bed) is formed by the posterior wall of the omental bursa and retroperitoneal structures between it and the posterior abdominal wall. Superiorly, the stomach bed includes part of the diaphragm (left crus), the spleen, the left suprarenal gland, and upper pole of the left kidney. Inferiorly, the stomach bed includes the body and tail of pancreas, transverse mesocolon, left colic flexure, the splenic artery, and, in some people, the transverse colon.
Anatomic regions. Anatomically, the stomach is subdivided arbitrarily into four regions: the fundus, body (corpus), cardia, and antrum (Fig. 12-3). It connects with the esophagus and duodenum through the cardia and pyloric canal, respectively. The fundus (superior part of the stomach) is a dome-shaped area that lies above an imaginary horizontal plane passing through the cardiac orifice. The fundus extends to the left and superiorly from the cardia region. The body lies between the fundus and the antrum. It is the largest part of the stomach and extends to the level of the incisura angularis on the lesser curve. Confusion can arise when fundus is used as a gross anatomic term including both the body and the fundus, and this use of the term should be avoided.13 The term “oxyntic mucosa” is a useful term to describe acid-producing mucosa. It is useful histologically when biopsies are clearly “oxyntic” but their precise site is unknown.
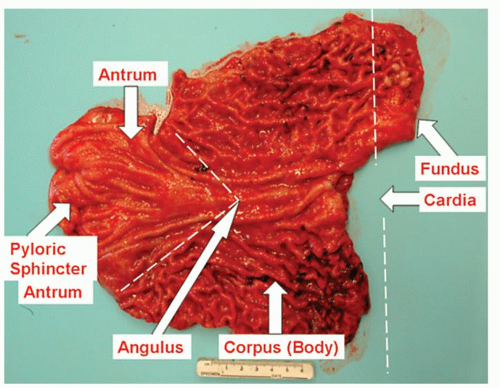 Figure 12-3. Areas of the stomach shown in this total gastrectomy specimen in a patient with an E-cadherin germline mutation but no gross abnormality. The pyloric sphincter is clearly identified on the left as a circular constriction. |
The antrum occupies the lower one-quarter to one-third of the stomach. Its approximate boundaries can be defined by a line drawn from the incisura angularis to the opposite wall; its junction on the greater curve is determined by the approximate ends of the gastric rugae (Fig. 12-3). When the stomach is distended with air at endoscopy to permit visualization, the antrum is designated as beginning on the greater curvature at the point where the gastric body folds end and where the smooth, “foldless” antrum begins. The opening into the duodenum is the pyloric orifice, and its position is usually indicated by a circular groove on the surface of the organ, termed the pyloric constriction, which indicates the position of the pyloric sphincter.
The pyloric sphincter is the most distal part of the antrum and controls the flow of gastric contents into the duodenum. The pyloric sphincter, similar to the lower esophageal sphincter, is several centimeters long (Fig. 12-3). Its narrow lumen, which passes between the antrum and the duodenum, is referred to as the pyloric canal. In endoscopic parlance, the term pylorus is commonly used interchangeably with pyloric canal and pyloric sphincter region. Some use the term pyloric antrum to designate the gastric antrum and pyloric canal to indicate the pyloric sphincter region. We prefer to avoid the prefix term pyloric in order to avoid confusion and to use the simpler terms antrum and pylorus.
The cardia region refers to a short zone measuring from a few millimeters to several centimeters where the stomach immediately adjoins the esophagus. It is so named because of the mucus-secreting glands (cardiac glands) it contains. When the term cardia region is used in clinical or macroscopic terms, it usually denotes the most proximal (juxtaesophageal) stomach. The gastric cardia starts at the gastroesophageal junction where the squamous mucosa terminates (in patients in whom Barrett’s esophagus is not a consideration). There is no consensus on the endoscopic landmarks for the gastric cardia. Western gastroenterologists arbitrarily define the distal extent of the cardia as the level of the most proximal gastric folds (Fig. 12-4).14 Japanese gastroenterologists define the proximal extent of the EGJ as the distal end of the lower esophageal palisade vessels.15, 16 These palisades are the termination of esophageal vessels that run longitudinally in the submucosal layer of the body of the esophagus, where their structure is truncal and consists of a few
large columns.16 At the cardia, the veins penetrate the muscularis mucosae and become superficial, forming the palisade vessels in the lamina propria beneath the epithelium. At the distal end of the palisade vessels, these veins merge with the submucosal venous network of the stomach. These two landmarks are similar but not always identical (Fig. 12-4).
large columns.16 At the cardia, the veins penetrate the muscularis mucosae and become superficial, forming the palisade vessels in the lamina propria beneath the epithelium. At the distal end of the palisade vessels, these veins merge with the submucosal venous network of the stomach. These two landmarks are similar but not always identical (Fig. 12-4).
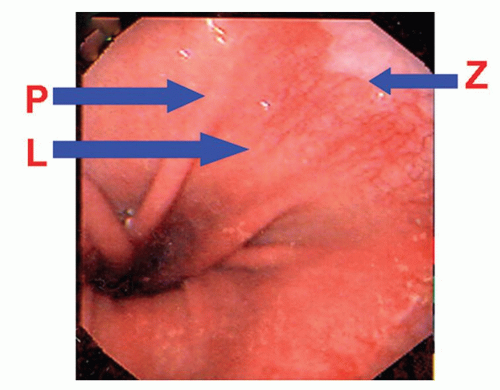 Figure 12-4. Endoscopic image of the cardia region. In Western countries, the cardia begins distally at the proximal end of the gastric longitudinal folds in the partially deflated stomach (P). In Japan, the lower end of the cardia is determined by the lower limit of palisade vessels (L). While both of these definitions are almost the same, as can be seen here, the palisade vessels actually extend a little distal to the upper end of the gastric folds. The upper end of the cardia is the squamocolumnar junction or Z-line (Z). (Image courtesy of Dr. Worth Boyce.) |
The squamocolumnar junction between the esophagus and the stomach is not an abrupt horizontal transition. Rather, there are grossly visible inter-digitating tongues referred to as the ora serrata or, more commonly in endoscopic circles, as the Z-line (Fig. 12-4).
As indicated subsequently in the section on histology, the histologic zones of the stomach do not correspond precisely to the gross anatomic regions. The mucosa in the body of the stomach, especially on the greater curvature, is thrown into numerous thick folds, or rugae, which run in a longitudinal direction. In the fundus, the lesser curvature of the body and, in the antrum, the mucosal folds commonly flatten when the stomach is distended with air or barium. With isotopically labeled meals, a midgastric transverse band can be identified in the food-filled stomach. There is no known anatomic correlate to explain its appearance.17 When the gastric body mucosa is viewed with a hand lens or close up at endoscopy, shallow slits or furrows, the areae gastricae, are seen. These represent the furrows between undulations of the mucosa that range up to 5 mm in diameter.
Blood vessels and lymphatics
Arterial Supply The stomach has a rich blood supply from all three branches of the celiac axis (left gastric, splenic, and common hepatic arteries. The lesser curvature receives blood from two sources. The lesser curvature of the stomach is supplied by the right gastric artery (that arises from the hepatic branch of the celiac near the pylorus) inferiorly and the left gastric artery (that arises directly from the celiac artery) superiorly, which also supplies the cardiac region. The greater curvature receives blood from two sources. The greater curvature is supplied by the right gastroepiploic artery (that originates from the gastroduodenal branch of the hepatic artery) inferiorly and the left gastroepiploic artery (which arises from the splenic artery) superiorly. The fundus of the stomach is supplied by short gastric branches from the splenic artery.
Venous Drainage The lesser curvature is drained by the right and left gastric veins, which run next to the arteries and drain into the portal vein. The fundus is supplied by short gastric veins that ultimately join the splenic vein, again finishing in the portal vein. Branches of the short gastric veins and the left coronary gastric vein at the cardia drain the lower esophagus. Blood from the lower portion of the greater curvature is drained by the right gastroepiploic vein, which enters the superior mesenteric vein and thence the portal vein.
Lymphatics The lymphatic drainage from the stomach generally follows the main arteries and is named accordingly. Beginning in the gastric mucosa, the lymphatics pass through the gastric wall and go toward one or the other of the curvatures, draining to four main areas, along the routes of the arteries and veins. The largest is for most of the lesser curvature and the lower end of the esophagus, along the left gastric artery to the left gastric nodes. The distal portion of the lesser curvature in the region of the pylorus drains to the right gastric nodes that drain to the hepatic nodes. The proximal part of the greater curvature drains along the gastroepiploic and splenic vessels, terminating in splenic nodes in the hilum of the spleen. The distal portion of the greater curvature drains to the right gastroepiploic nodes in the greater omentum and to pyloric nodes at the head of the pancreas. Ultimately, the branches from all of these groups drain into the celiac nodes located around the celiac trunk as it arises from the abdominal aorta. From the celiac nodes, lymph drains directly into the thoracic duct. It is important to understand gastric lymphatic drainage as veins provide routes for spread of gastric cancer, when present, directly to the liver, while from the celiac nodes, the thoracic duct provides direct access to the systemic circulation.
Nerve supply. The parasympathetic nerve supply is from the vagus nerve and its branches. The right vagus nerve enters the abdomen as one or two trunks on the posterior side of the esophagogastric junction. It supplies branches to both surfaces of the stomach. The left vagus nerve also has one or two trunks, and it enters the abdomen on the anterior surface of the stomach, where it is present because of the rotation of the stomach early in its development (see “Embryology” section above). Shortly after entering the abdomen, the anterior vagal nerve gives off a hepatic branch and the posterior trunk gives off a celiac branch. Thus, transection of these trunks below these branches only results in gastric denervation. Even more selective denervation of the fundic gland mucosa (highly selective vagotomy) can be achieved by preserving the terminal portions of the vagal nerves that supply the gastric antrum. Vagal nerve fibers connect with ganglion cells located between the circular and longitudinal fibers of the muscularis propria (Auerbach’s plexus) and with submucosal ganglion cells (Meissner’s plexus). From these plexi, postganglionic fibers innervate both glands and muscle.
The sympathetic nerve supply to the stomach is from the celiac plexus via branches that follow the gastric and gastroepiploic arteries. There are also sympathetic branches from the right and left phrenic nerves. These nerves contain afferent pain fibers as well as motor fibers to the pyloric sphincter region. Vagal stimulation from ghrelin mediated pituitary stimulation produces release of ghrelin and orexin that stimulates gastric secretion prior to food ingestion, the former resulting in increased gastric acid secretion via acetyl choline, and the latter that also stimulates a craving for food.
Histology
This section focuses on the light microscopic appearance of the stomach. Comprehensive reviews of the electron microscopic appearance are available.18, 19, 20 All parts of the stomach have the same basic structural layers (mucosa, submucosa, muscularis propria, subserosa, and serosa).
Mucosa. Traditionally, but for no good reason that we are aware of, the term crypt (crypts of Lieberkuhn) is applied to the small and large intestine, but in the stomach, they are pits. We are willing to accept either terminology but will use “pit” as terms such as “pit pattern” are now generally accepted. Gastric pits are divided into three histologic zones: superficial zone (surface and pit epithelium); neck zone, which is the regenerative region of the stomach; and deep or glandular zone. While the surface and neck zone cells are uniform throughout the stomach, the underlying glands differ in structure and function by region, and the histologic zones of the stomach are classified according to these types of glands: cardiac, oxyntic, antral.
At the microscopic level, this esophagogastric transition zone frequently contains alternating islands of squamous and columnar epithelium. Similarly, in the pyloric sphincter region, there may be short segments containing a blend of gastric columnar and small intestinal columnar epithelial cells.
The Superficial Zone (Surface and Pit Epithelium) The gastric mucosa is covered by tall (20-40 µm) columnar mucous cells that are invaginated to form the pits or foveolae. The surface epithelial cells are replaced every 4 to 6 days.21 The mucus in the surface epithelial cells occupies the luminal part of the cells (Fig. 12-5), occupying up to about 80% of the cell. Nuclei are regularly oriented and are normally located in the basal part of the cell, and no more than one-fifth of the distance toward the lumen; when reactive changes are present, the
mucin is reduced and the nuclei occupy correspondingly more of the cell (see Chapter 13). Mucous cells lining the gastric lumen secrete mucus as well as bicarbonate ions. The bases of these pits are in direct continuity with the underlying glands. The surface epithelial cells appear similar to each other in all gland zones except those in the cardia region, where they may be taller and narrower. Occasional intraepithelial lymphocytes are normally present. Sometimes, the surface configuration of the mucosa of the antral and cardiac gland mucosa has a villous configuration. This appearance is a normal variant and should not be confused with intestinal metaplasia of the stomach, because the epithelial cells are normal, not metaplastic. The entire superficial zone of the stomach including the superficial part of the mucous secreting glands are immunoreactive to MUC5, and the deeper parts with MUC6, a staining pattern also seen in antral mucosa, Table 12-1, and illustrated in Chapter 14. These cells are strongly diastase PAS positive but do not stain with Alcian blue except in the cardia.
mucin is reduced and the nuclei occupy correspondingly more of the cell (see Chapter 13). Mucous cells lining the gastric lumen secrete mucus as well as bicarbonate ions. The bases of these pits are in direct continuity with the underlying glands. The surface epithelial cells appear similar to each other in all gland zones except those in the cardia region, where they may be taller and narrower. Occasional intraepithelial lymphocytes are normally present. Sometimes, the surface configuration of the mucosa of the antral and cardiac gland mucosa has a villous configuration. This appearance is a normal variant and should not be confused with intestinal metaplasia of the stomach, because the epithelial cells are normal, not metaplastic. The entire superficial zone of the stomach including the superficial part of the mucous secreting glands are immunoreactive to MUC5, and the deeper parts with MUC6, a staining pattern also seen in antral mucosa, Table 12-1, and illustrated in Chapter 14. These cells are strongly diastase PAS positive but do not stain with Alcian blue except in the cardia.
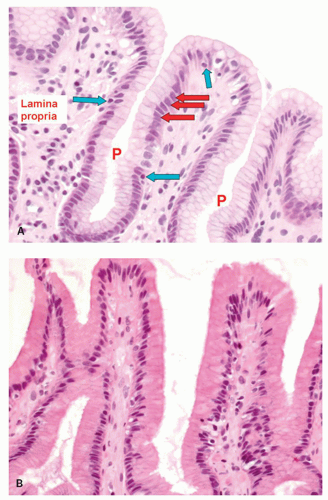 Figure 12-5. A: In the gastric pits (P), superficial cells in the stomach are mucin-producing throughout the entire stomach. Each nucleus has an apical mucin vacuole that usually occupies well over 50% of the cell (red arrows). Occasional intraepithelial lymphocytes are also visible (blue arrows). Depending on the staining technique employed, the mucous cells maybe almost clear as in (A), or eosinophilic as in (B). |
Table 12-1 Endocrine Cells of the Stomach | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The Neck Zone (Middle Zone) The neck zone (middle zone) has mucous cells as well as immature stem cells. This is the generative zone for the stomach, and all cells above and below are derived from this region. Mitotic figures are surprisingly sparse, although stains for Ki-67 show virtually all of the cells to be in the proliferative phase (Fig. 12-6). The immature stem cells migrate upward to renew the surface epithelium or downward to form the differentiated cells of the glands. These cells are also PAS positive.
The Deep (Glandular) Zone Glands are found in the deep zone. In contrast to surface mucous cell that is continuous over the entire stomach surface, regional differences are mainly in the composition of gastric tubular glands within each compartment.
Cardiac Gland Mucosa The gastric cardia has branched tubular glands that are primarily mucous-type glands, oxyntic-type glands, or both.22, 23 Those that are mucous but clearly have residual specialized elements are called cardio-oxyntic. Unlike the antrum, in which the deep glands are diffuse, in the cardia, they often are compartmentalized, akin to lobules, which in normal stomachs allow ease of identification at low power. Mucus-secreting glands are similar in appearance on conventional stains to those of the gastric antrum and Brunner’s glands, except for the compartmentalization and lack of overt endocrine cells so readily seen in the gastric antrum (Fig. 12-7). In contrast to antral glands, portions of the cardiac glands may contain sialomucins that stain with Alcian blue at pH 2.5. Parietal cells are commonly scattered within the cardiac glands, although endocrine and chef cells are rare. Mucous cell-lined cysts are a frequent finding
(Fig. 12-7). The pits in the cardiac region are very shallow, occupying less than one-quarter of the overall mucosal thickness (Fig. 12-8)—ranging from 0.5 to 1.5 mm—and are immunoreactive with MUC6. There are at best rare endocrine cells in the gastric cardia—there are no data to suggest that they have physiologic or pathologic significance.
(Fig. 12-7). The pits in the cardiac region are very shallow, occupying less than one-quarter of the overall mucosal thickness (Fig. 12-8)—ranging from 0.5 to 1.5 mm—and are immunoreactive with MUC6. There are at best rare endocrine cells in the gastric cardia—there are no data to suggest that they have physiologic or pathologic significance.
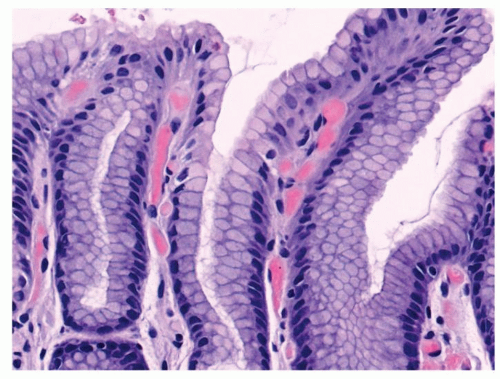 Figure 12-6. Mucous neck region stained with Mib-1 to show the marked proliferation of this region. The paradox is how inconspicuous mitotic figures are in this region. |
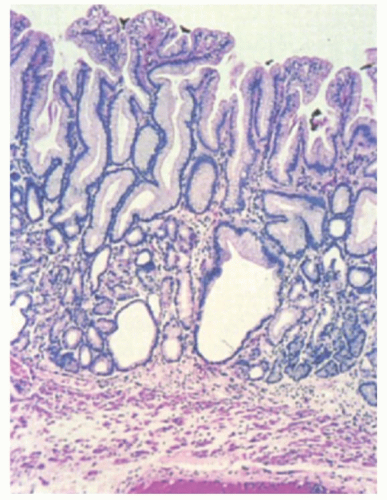 Figure 12-7. Cardiac gland mucosa. The glands in the bottom half of the section are a mix of mucous glands (cardiac glands) and fundic gland elements. Mucosal cysts, as seen here, are a common finding in cardiac gland mucosa. Note the vague lobularity that is very characteristic of cardia. Occasionally, pancreatic-type cells can be seen in these glands but appear to be without significance. |
Occasionally, pancreatic-type glands can be seen in the cardia in the form of exocrine cells that are immunoreactive with lipase or trypsin. They can be admixed with simple mucous-producing cells or oxyntocardiac mucosa with parietal and mucous-producing glands. Chief cells appear to be rare in the cardia. Endocrine components admixed with heterotopic pancreas are vanishingly rare, and endocrine neoplasms even more so. Also, at the squamocolumnar junction, the so-called multilayered epithelium can be seen (Fig. 12-9), which seems to be associated with the development of Barrett’s esophagus. They are sometimes ciliated. They are discussed in more detail in Chapter 10.
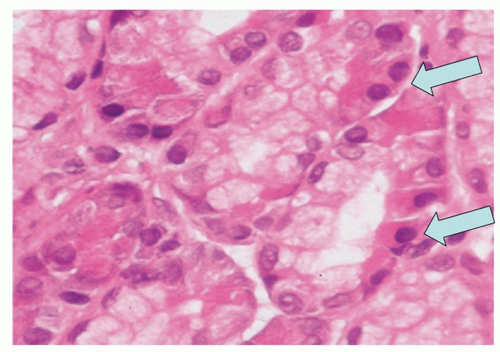 Figure 12-8. Cardiac glands with pancreatic-type cells (arrows). |
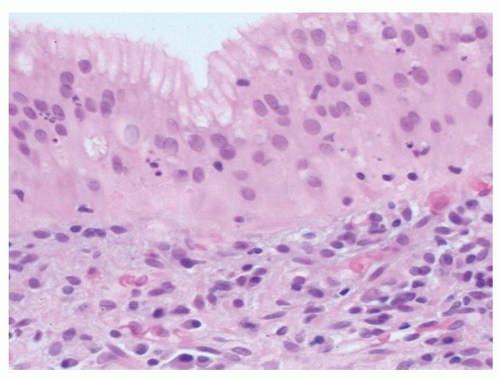 Figure 12-9. Transitional mucosa (mutilayered epithelium) that is sometimes seen in the cardiac region. The most superficial cells are mucous secreting, but beneath that, the cells and their nuclei are stratified as seen in squamous mucosa of the esophagus. These are discussed further in Chapter 10. |
Oxyntic Mucosa (Fundic and Corpus Gland Mucosa) Oxyntic gland literally means acid producing so is a good name for normal corpus and fundic gland mucosa. Surprisingly, the thickness of the oxyntic mucosa varies. It is thickest in the greater curve, gradually thins as the antrum is approached, and is thinnest on the lesser curve (Fig. 12-10).
Related posts:
 Disorders of Endocrine Cells
Disorders of Endocrine Cells
 Gastrointestinal Manifestations of Extraintestinal Disorders and Systemic Disease
Gastrointestinal Manifestations of Extraintestinal Disorders and Systemic Disease
 Inflammatory Disorders of the Esophagus: Reflux and Nonreflux Types
Inflammatory Disorders of the Esophagus: Reflux and Nonreflux Types
 Small Bowel Mucosal Disease
Small Bowel Mucosal Disease
 Stomach and Proximal Duodenum: Inflammatory and Miscellaneous Disorders
Stomach and Proximal Duodenum: Inflammatory and Miscellaneous Disorders
 Small and Large Bowel Polyps and Tumors
Small and Large Bowel Polyps and Tumors
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


